Дейтерий и эволюция
Новая статья - 2010 год
Рисунок на обложке. Гипотеза “Большого взрыва” предполагает образование Вселенной более десяти миллиардов лет тому назад из бесконечно малой точки со сверхкритической плотностью.
О.В. Мосин

ДЕЙТЕРИЙ, ТЯЖЁЛАЯ ВОДА, ЭВОЛЮЦИЯ И ЖИЗНЬ
Вселенная, образовавшаяся в результате “Большого взрыва” несколько десятков миллиардов лет тому назад, была значительно горячее и плотнее, чем сейчас и состояла, в основном, из одного элемента – водорода. Дейтерий сформировался в последующие мгновения эволюции Вселенной в результате столкновения свободного нейтрона и протона при температурах миллион градусов Цельсия. А ещё позже два атома дейтерия вошли в состав ядра гелия, который состоит из двух протонов и двух нейтронов. Таким образом, дейтерий может служить своеобразным индикатором эволюции Вселенной, поскольку количество дейтерия в мире постоянно. Вплоть до настоящего времени считалось, что в процессе формирования гелия израсходовался почти все атомы дейтерий, и лишь 10 тысяч атомов дейтерия остались неизрасходованными. Исходя из этого количества дейтерия в мире, природная распространённость дейтерия составляла по расчётам исследователей не более 0.015% (от общего числа всех атомов водорода). Однако, недавно было показано, что это не так – что дейтерия во Вселенной гораздо больше. Статья знакомит читателя с эволюцией Вселенной, образованием дейтерия и его ролью в молекулярной эволюции.
Началось всё с того, что проводя наблюдения Млечного Пути, американские учёные обнаружили что дейтерия — тяжёлого водорода – содержится в нём значительно больше, чем об этом свидетельствовали данные предыдущих исследований. По мнению астронома Джеффри Лински (Jeffrey L. Linsky) из университета Колорадо (University of Colorado), руководившего исследованием, эта новая информация может радикальным образом изменить теоретические положения о формировании звёзд и галактик.
Тяжёлый водород "прятался" от телескопов за скоплениями межзвёздной пыли и часто был недоступен для наблюдений в силу своей непрозрачности. Астрономы использовали данные ультрафиолетового телескопа FUSE (Far Ultraviolet Spectroscopic Explorer). Дейтерий создаёт характерное свечение в ультрафиолетовом диапазоне, благодаря которому разглядеть тяжёлый водород удалось именно с помощью FUSE.
До настоящего времени считалось, что природная распространённость дейтерия составляет не более 0.015% (от общего числа всех атомов водорода). Это количество зависит как от природы вещества, так и от общего количества материи, сформированной в ходе эволюции Вселенной. Теперь очевидно, что дейтерия в природе намного больше, чем предполагалось раннее.
Но с чем это может быть связано? Источником дейтерия во Вселенной являются вспышки сверхновых и термоядерные процессы, идущие внутри звёзд. Возможно этим объясняется тот факт, что мировое количество дейтерия повышается в период глобальных потеплений и изменений климата. Однако дейтерий довольно быстро разрушается в этих звёздах.
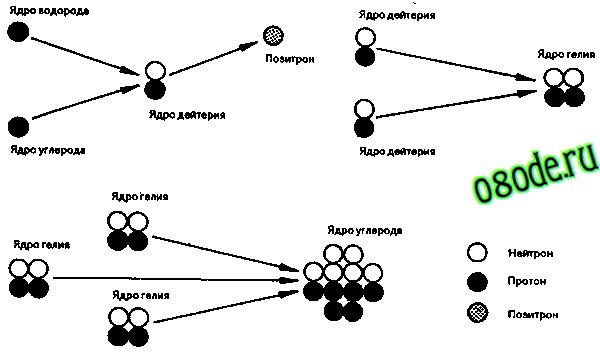
Рис. 1. Возможные ядерные реакции с участием дейтерия
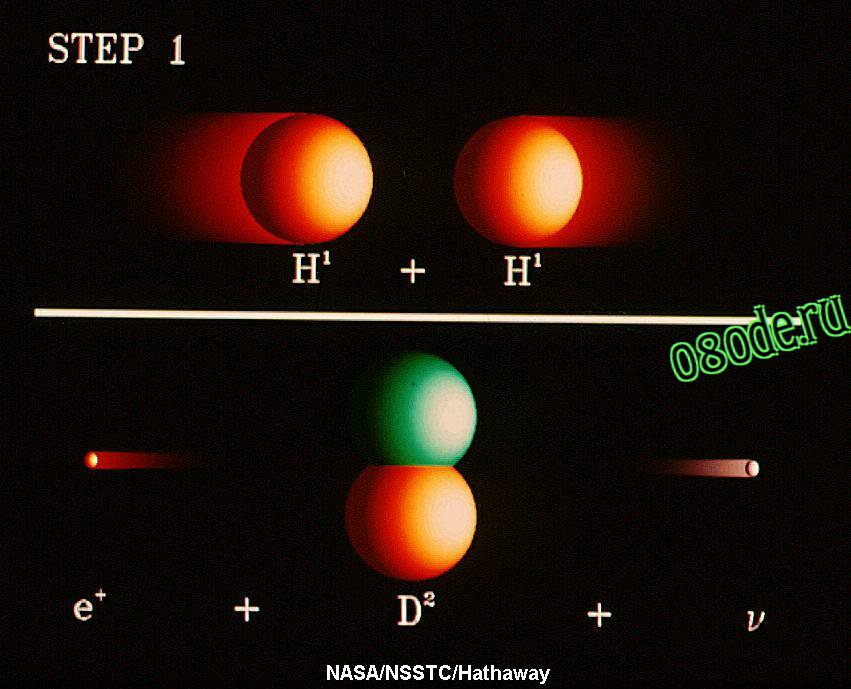
Рис. 2. Первый этап эволюции Вселенной. Образование дейтерия из двух атомов водорода
Дело в том, что наряду с водородом в первые мгновения после Большого взрыва образовалось и огромное количество его изотопа дейтерия. Исходя из предыдущих наблюдений, учёные постановили, что больше трети первоначально образованного дейтерия потратилось на создание звёзд. Однако, оказывается, что дейтерия в Млечном Пути намного больше, чем предполагали ранее. В частности, на звездообразование потрачена не треть, а всего 15% изотопа и он распределён неравномерно. Эти данные могут говорить о том, что для формирования звёзд требовалось значительно меньше водорода, превратившегося затем в гелий. Так же это может оказаться существенным основанием для пересмотра теории эволюции галактик и звёзд.
Если это так, то необходимо также пересмотреть теорию молекулярной эволюции и эволюции жизни на нашей планете, поскольку жизнь напрямую связана с водой и зарождалась в ней. Но была ли это обычная вода? Ещё 10 лет тому назад автор этой статьи, будучи аспирантом Московской государственной академии тонкой химической технологии им. М. В. Ломоносова в группе академика РАМН В. И. Швеца выдвинул предположение, что первичный “первобытный бульон”, в котором зарождалась жизнь в виде первых коорцерватов, был насыщен тяжёлой водой вследствии того, что в атмосфере Земли не было защитного озонового слоя и вулканические геотермальные и электрические процессы в горячей атмосфере, насыщенной водой могли привести к обогащению гидросферы тяжёлой водой. Но тогда мало кто из учёных увлёкся этой идеей, хоть и напрямую никто не отвёрг её. И только сейчас стало очевидным, что исследователи пренебрегали дейтерием в своих расчётах.
Астрономы считают, что гравитационное поле Земли – недостаточно сильно для удержания водорода, и наша планета постепенно теряет водород в результате его диссоциации в межпланетное пространство. Водород улетучивается быстрее тяжелого дейтерия, который способен накапливаться. Так что в течение всей эволюции должно происходить накопление дейтерия в атмосфере и в поверхностных водах.
Если это так, то необходимо пересмотреть эволюцию всего живого на нашей планете, чтобы смоделировать и предсказать дейтерированные формы жизни. Тем более, что их можно легко создать в современных условиях – макромолекулы ДНК, белков, липидов и сахаров – вот те главные компоненты для конструирования дейтерированных мембран и изучения гидрофобных взаимодействий между дейтерированными молекулами.
Отдельный вопрос – генетика дейтерированных клеток и изучение распределения наследственного аппарата, а также физиология, цитология и морфология клетки при росте на тяжёлой воде.
Модели дейтерированных систем довольно легко прогнозировать и конструировать в лабораторных условиях. Нами были получены адаптированные к тяжёлой воде штаммы бактерий, относящиеся к различным таксономическим группам. арактерной особенностью объектов являлось то, что весь биологический материал клетки вместо природного водорода содержал дейтерий.
Дейтерированные клетки адаптированных к максимальной концентрации тяжёлой воды в среде – весьма удобные объекты для исследования. В процессе роста клеток на тяжёлой воде в них синтезируются макромолекулы, в которых атомы водорода в углеродном скелете полностью замещены на дейтерий. Такие дейтерированные макромолекулы претерпевают структурно-адаптационные модификации, необходимые для нормального функционирования клетки в тяжёлой воде. Но эти изменения не единственны; физиология, морфология, цитология клетки, а также генетический аппарат клетки также подвергается воздействию и модификации в тяжёлой воде.
Способность микроорганизмов расти в искусственных условиях на тяжёловодородных средах, в которых все атомы протия заменены на дейтерий является одним из интереснейших биологических феноменов (О.В. Мосин, Д.А. Складнев, В. И. Швец, 1996).
Тяжёлая вода (оксид дейтерия) — имеет ту же химическую формулу, что и обычная вода, но вместо атомов водорода содержит два тяжёлых изотопа водорода — атомы дейтерия. Формула тяжёловодородной воды обычно записывается как: D2O или 2H2O. Внешне тяжёлая вода выглядит как обычная — бесцветная жидкость без вкуса и запаха.
В природных водах соотношение между тяжёлой и обычной водой составляет 1:5500 (в предположении, что весь дейтерий находится в виде тяжёлой воды D2O, хотя на самом деле он частично находится в составе полутяжёлой воды HDO).
Тяжёлая вода токсична в высоких концентрациях, химические реакции в её среде проходят несколько медленнее, по сравнению с обычной водой, водородные связи с участием дейтерия несколько сильнее обычных. Эксперименты над млекопитающими показали, что замещение 25% водорода в тканях дейтерием приводит к стерильности, более высокие концентрации приводят к быстрой гибели животного. Однако некоторые микроорганизмы способны жить в 70%-ной тяжёлой воде) (простейшие) и даже в чистой тяжёлой воде (бактерии). Человек может без видимого вреда для здоровья выпить стакан тяжёлой воды, весь дейтерий будет выведен из организма через несколько дней. В этом отношении тяжёлая вода менее токсична, чем, например, поваренная соль.
Тяжёлая вода накапливается в остатке электролита при многократном электролизе воды. На открытом воздухе тяжёлая вода быстро поглощает пары обычной воды, поэтому можно сказать, что она гигроскопична. Производство тяжёлой воды очень энергоёмко, поэтому её стоимость довольно высока (ориентировочно 200-250 долларов за кг).
Физические свойства обычной и тяжёлой воды
Физические свойства
D2O
H2O
Молекулярная масса
20
18
Плотность при 20 C (г/см3)
1,1050
0,9982
t кристаллизации ( C)
3,8
0
t кипения ( C)
101,4
100
Важнейшим свойством тяжёлой воды является то, что она практически не поглощает нейтроны, поэтому используется в ядерных реакторах для торможения нейтронов и в качестве теплоносителя. Она используется также в качестве изотопного индикатора в химии и биологии. В физике элементарных частиц тяжёлая вода используется для детектирования нейтрино; так, крупнейший детектор солнечных нейтрино в Канаде содержит 1 килотонну тяжёлой воды.
Существует также и полутяжёлая (или дейтериевая) вода, у которой только один атом водорода замещен дейтерием. Формулу такой воды записывают так: DHO.
Термин тяжёлая вода применяют также по отношению к воде, у которой любой из атомов заменен тяжёлым изотопом:
- к тяжёлокислородной воде (в ней лёгкий изотоп кислорода 16O замещен тяжёлыми изотопами 17O или 18O).
- к тритиевой и сверхтяжёлой воде (содержащей вместо атомов 1H его радиоактивный изотоп тритий 3H).
Открытие тяжелой воды послужило толчком к выяснению фракционного состава воды. Вскоре была обнаружена сверхтяжелая вода Т20. В ее составе место водорода занимает его природный изотоп, еще более тяжелый, чем дейтерий. Это тритий (Т), он радиоактивен, атомная масса его равна 3. Тритий зарождается в высоких слоях атмосферы, где идут природные ядерные реакции. Он является одним из продуктов бомбардировки атомов азота нейтронами космического излучения. Ежеминутно на каждый квадратный сантиметр земной поверхности падают 8-9 атомов трития.
В небольших количествах сверхтяжелая (тритиевая) вода попадает на Землю в составе осадков. Во всей гидросфере одновременно насчитывается лишь около 20 кг Т20. Тритиевая вода распределена неравномерно: в материковых водоемах ее больше, чем в океанах; в полярных океанских водах ее больше, чем в экваториальных. По своим свойствам сверхтяжелая вода еще заметнее отличается от обычной: кипит при 104°С, замерзает при 4...9°С, имеет плотность 1,33 г/см3. Сверхтяжелую воду применяют в термоядерных реакциях. Она удобнее дейтериевой, так как чувствительнее в определении.
Перечень изотопов водорода не кончается тритием. Искусственно получены и более тяжелые изотопы4H и 5H,тожерадиоактивные.
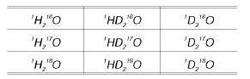
Если подсчитать все возможные различные соединения с общей формулой Н2О, то общее количество возможных «тяжёлых вод» достигнет 48. Из них 39 вариантов — радиоактивные, а стабильных вариантов всего девять:
Н2 6O, Н217O, Н218O, HD16O, HD17O, HD18O, D216O, D217O, D218O.
Таким образом, возможно существование молекул воды, в которых содержатся любые из пяти водородных изотопов в любом сочетании.
Этим не исчерпывается сложность изотопного состава воды. Существуют также изотопы кислорода. В периодической системе химических элементов Д.И. Менделеева значится всем известный кислород 16O. Существуют еще два природных изотопа кислорода – 17O и 18O. В природных водах в среднем на каждые 10 тысяч атомов изотопа 16O приходится 4 атома изотопа 17O и 20 атомов изотопа 18O.
По физическим свойствам тяжелокислородная вода меньше отличается от обычной, чем тяжеловодородная. Получают ее в основном перегонкой природной воды и используют как источник препаратов с меченым кислородом.
Помимо природных, существуют и шесть искусственно созданных изотопов кислорода. Как и искусственные изотопы водорода, они недолговечны и радиоактивны. Из них: 13O, 14O и 15O – легкие, 19O и 20O – тяжелые, а сверхтяжелый изотоп – 24O получен в 1970 году.
Существование пяти водородных и девяти кислородных изотопов говорит о том, что изотопных разновидностей воды может быть 135.
Основную же массу природной воды – свыше 99% – составляет протиевая вода – 1H216O. Тяжелокислородных вод намного меньше: 1H218O – десятые доли процента.
1H217O – сотые доли от общего количества природных вод. Только миллионные доли процента составляет тяжелая вода D2O, зато в форме 1HDO тяжелой воды в природных водах содержится уже заметное количество. Еще реже, чем D2O, встречаются и девять радиоактивных естественных видов воды, содержащих тритий:

Классической водой следует считать протиевую воду 1H216O в чистом виде, то есть без малейших примесей остальных 134 изотопных разновидностей. И хотя содержание протиевой воды в природе значительно превосходит содержание всех остальных вместе взятых видов, чистой 1H216O в естественных условиях не существует. Во всем мире такую воду можно отыскать лишь в немногих специальных лабораториях. Ее получают очень сложным путем и хранят с величайшими предосторожностями. Для получения чистой 1H216O ведут очень тонкую, многостадийную очистку природных вод или синтезируют воду из исходных элементов 1H2 и 16O, которые предварительно тщательно очищают от изотопных примесей. Такую воду применяют в экспериментах и процессах, требующих исключительной чистоты химических реактивов.
Но самые большие изотопные эффекты наблюдается для пар протий/дейтерий и протий/тритий. Двукратным увеличением массы дейтерона относительно протона и обуславливаются так называемые изотопные эффекты тяжёлой воды - энергия связи, константа диссоциации, подвижность, длина связи и т.д.
С первых экспериментов американца Креспи и Даболла в 1940-х годах прошлого века, вплоть до конца 90-х годов установилось устойчивое представление, что тяжёлая вода несовместима с жизнью и что высокие концентрации тяжёлой воды могут приводить к ингибированию многих жизненно-важных мутаций, включая блокировку митоза в стадии профазы, и даже в некоторых случаях вызывать спонтанные мутации.
Клетки животных способны выдерживать до 25-30% тяжёлой воды в среде, растений (50%), а клетки простейших микроорганизмов способны жить на 80% тяжелой воде. Однако, потом было доказано, что многие организмы могут быть адаптированы к росту на тяжёлой воде.
Тяжёлая вода высокой концентрации токсична для организма; химические реакции в её среде проходят несколько медленнее, по сравнению с обычной водой, водородные связи с участием дейтерия несколько сильнее обычных.
Тем не менее тяжелая вода играет значительную роль в различных биологических процессах. Систематическое изучение ее воздействия на животных и растения начато сравнительно недавно. Различные исследователи независимо друг от друга установили, что тяжелая вода действует отрицательно на жизненные функции организмов; это происходит даже при использовании обычной природной воды с повышенным содержанием тяжелой воды (рис. 3).
Влияние концентрации дейтерия на рост высших растений
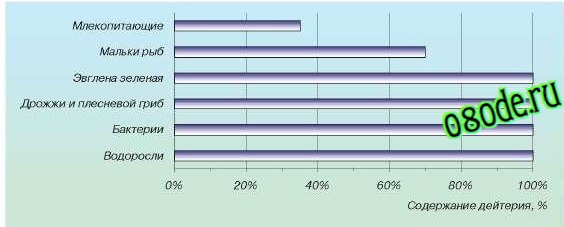
Рис.3. Выживаемость различных организмов в воде с различными концентрациями дейтерия
Подопытных животных поили водой, 1/3 часть которой была заменена водой состава HDO. Через недолгое время начиналось расстройство обмена веществ животных, разрушались почки. При увеличении доли тяжелой воды животные погибали.
На развитие высших растений тяжелая вода также действует угнетающе; если их поливать водой, на половину состоящей из тяжелой воды, рост прекращается (рис. 4).
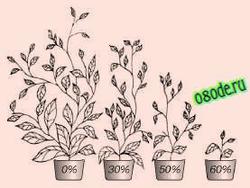
Рис 4. Поливка помидорной рассады 30, 50 и 60%-ной тяжёлой водой ингибирует рост растения (по данным Креспи и Катца, 1972).
При попадании клеток в дейтерированную тяжёловодородную среду из них не только исчезает протонированная вода за счет реакции обмена Н2О-D2О, но и происходит быстрый H±D обмен в гидроксильных, сульфгидрильных и аминогруппах всех органических соединений, включая белки, нуклеиновые кислоты, липиды, сахара. Только С—Н-связь не подвергается обмену и соединения типа С—D синтезируются «de поvo».
Интересно, что после обмена H±D ферменты не прекращают своей функции (Themson et al., 1966; Денько, 1974), но изменения в результате изотопного замещения за счет первичного и вторичного изотопных эффектов (Thomson, 1963; Halevy, 1963), а также действие тяжёлой воды как растворителя (большая структурированность и вязкость по сравнению с обычной водой) приводят к изменению скоростей (замедлению) и специфичности ферментативных реакций в тяжёлой воде.
Присутствие дейтерия в биологических системах приводит к изменениям структуры и свойствам жизненно-важных макромолекул таких как дезоксирибонуклеиновые кислоты (ДНК) и белки. При этом различают первичные и вторичные изотопные эффекты дейтерия в зависимости от того, какое положение занимает атом дейтерия в молекуле. Наиболее важными для структуры макромолекулы связи являются динамические короткоживущие водородные (дейтериевые) связи. Они формируются между соседними атомами дейтерия (водорода) и гетероатомами кислорода, углерода, азота, серы и т.д. и играют главную роль в поддержании пространственной структуры макромолекулярных цепей и как эти структуры взаимодействуют с другими соседними макромолекулярными структурами, а также с тяжелой водной окружающей среды.
Структурно-динамические свойства клеточной мембраны, которые в большинстве зависят от качественного и количественного состава липидов, также могут изменяться в присутствии тяжёлой воды. Полученный результат объясняется тем, что клеточная мембрана является одной из первых органелл клетки, которая испытывает воздействие тяжёлой воды, и тем самым компенсирует реалогические параметры мембраны (вязкость, текучесть, структурированность) изменением количественного и качественного состава липидов.
В клетках бактерий одним из важнейших инструментов регу ляции метаболизма является мембрана, объединяющая в себе аппараты биосинтеза полисахаридов, трансформации энергии, снабжении клетки метаболитами и участвующая в биосинтезе белков, нуклеиновых кислот и липидов (Гельман с соавт., 1972; Коротяев, 1973; Островский, 1973). Можно предположить, что в адаптации к тяжёлой воде мембраны играют не последнюю роль. Однако до сих пор не по нятно, что происходит с мембранами, как они реагируют на замену сре ды и какое это имеет значение для выживания клеток на среде, лишен ной протонов. Структурно-динамические свойства клеточной мембраны, которые в большинстве зависят от качественного и количественного состава липидов, также могут изменяться в присутствии тяжёлой воды. Так, сравнительный анализ липидного состава дейтерированных клеток B. subtilis, полученных при росте на тяжёлой воде показал различия в количественном составе мембранных липидов по сравнению с обычной водой (рис. 5). Примечательно, что в образце полученном с тяжёлой воды соединения, имеющие времена удерживания - 33.38; 33.74 и 33.2 мин не детектируются (рисунок 5 б). Полученный результат, по видимому, объясняется тем, что клеточная мембрана является одной из первых органелл клетки, которая испытывает воздействие тяжёлой воды, и тем самым компенсирует реалогические параметры мембраны (вязкость, текучесть, структурированность) изменением количественного состава липидов.
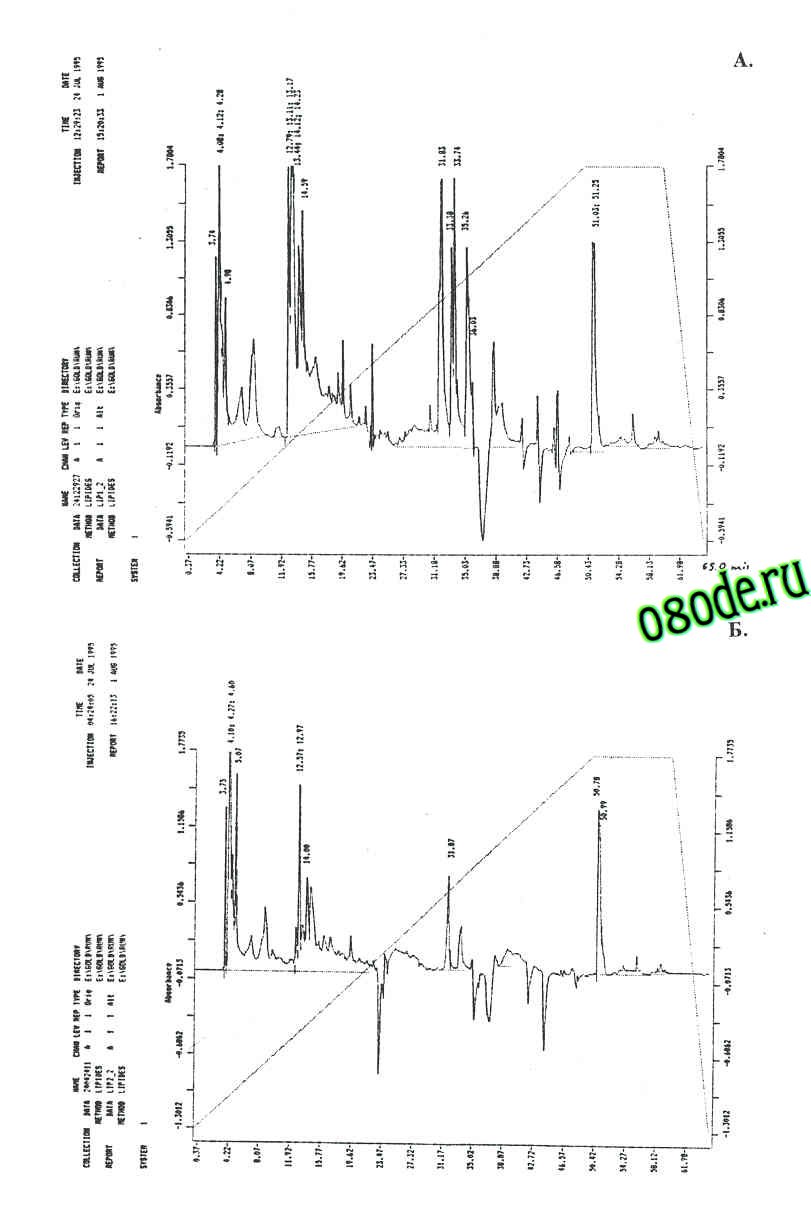
Рис.5. Липидные профили протонированных (а) и дейтерированных (б) клеток Basilus subtilis; хроматограф Beckman Gold System (США), снабжённый насосом Model 166 (США) и детектором Model 126 (США); неподвижная фаза: Ultrasphere ODS 5 мкм; 4.6 x 250 мм; подвижная фаза: линейный градиент 5 мМ KH2PO4-ацетонитрил; 100% в течении 50 мин; скорость подачи: 0.5 мл/мин; детекция при 210 нм.
Также возможно эффекты, наблюдаемые при адаптации к тяжёлой воде связаны с образованием в тяжёлой воде конформаций молекул с иными структурно-динамическими свойствами, чем конформаций, образованных с участием водорода, и поэтому имеющих другую активность и биологические свойства. Так, по теории абсолютных скоростей разрыв СH-связей может происходить быстрее, чем СD-связей, подвижность иона D+ меньше, чем подвижность Н+, константа ионизации тяжёлой воды меньше константы ионизации обычной воды. Всё это отражается на кинетике химической связи и скорости хим. реакций в тяжёлой воде.
Связи, образованные атомами углерода с дейтерием немного прочнее, чем СН-связи из-за того, что частота колебания дейтерона, имеющего большую массу (в два раза большую, чем протон) и размер меньше частоты колебания протона и тем самым, это стабилизирует связь.
Другое важное свойство определяется самой пространственной структурой тяжёлой воды, которая имеет тенденцию сближать гидрофобные группы макромолекулы, чтобы минимизировать их эффект на водородную (дейтериевую) связь в присутствии молекул тяжёлой воды. Так что структура спирали, каковой является ДНК в присутствии тяжёлой воды стабилизируется. Кроме этого, отмечены радиопротекторные свойства тяжёлой воды на клетки печени обезьяны, в которой экспонировались эти клетки. Также было показано, что жизненный цикл плоских червей, выращенных на тяжёлой воде увеличивается в 1.5 раза по-сравнению с червями, выращенными на обычной воде (М.Шепенинов, 2006).
Вероятно, клетка реализует лабильные адаптивные механизмы, которые способствуют функциональной реорганизации работы жизненно-важных систем в тяжёлой воде. Так, например, нормальному биосинтезу и функционированию в тяжёлой воде таких биологически активных соединений, как нуклеиновые кислоты и белки способствует поддержание их структуры посредством формирования водородных (дейтериевых) связей в молекулах.
Связи, сформированные атомами дейтерия различаются по прочности и энергии от аналогичных водородных связей. Различия в нуклеарной массе атома водорода и дейтерия косвенно могут служить причиной различий в синтезах нуклеиновых кислот, которые могут приводить в свою очередь к структурным различиям и, следовательно, к функциональным изменениям в клетке.
Ферментативные функции и структура синтезируемых белков также изменяются при росте клеток на тяжёлой воде, что может отразиться на процессах метаболизма и деления клетки.
Способность к адаптации в высоких концентрациях тяжёлой воды связана с эволюционным уровнем организации, т. е. чем ниже уровень развития живого, тем выше способность к адаптации (О.В. Мосин, Д.А. Складнев, В.И. Швец, 1996).
Изменения соотношения основных метаболитов в процессе адаптации к тяжеловодородной среде также может являться причинами гибели клеток. Клетки высших организмов погибают при содержании тяжёлой воды в составе тела свыше 30%, но микроорганизмы, легко приспосабливающиеся к резким изменениям среды обитания, способны жить и размножаться даже в 98%-ной тяжёлой воды (Мосин О.В, 1996).
Давно замечено, что адаптация к тяжёлой воде проходит легче при постепенном увеличении содержания дейтерия в среде (Денько Е.И, 1970), так как чувствительность к тяжёлой воде разных ключевых систем различна. Практически даже высокодейтерированные среды содержат протоны от 0,2—10%. Возможно, что остаточные протоны в момент адаптации к тяжёлой воде облегчают перестройку к изменившимся условиям, встраиваясь именно в те участки, которые наиболее чувствительны к замене. Если это так, то встраивание протонов должно приводить к накоплению легкого изотопа в органическом материале клеток и соответственно к обогащению тяжелым изотопом среды культивирования.
Адаптация к тяжёлой воде является фенотипическим феноменом, поскольку адаптированные к тяжёлой воде клетки возвращаются к нормальному росту в протонированных средах после некоторого лаг-периода (О.В. Мосин с соавт., 1996). В то же время обратимость роста на Dи Н-средах не исключает возможности изменения метаболизма дейтерированных клеток, т.е. морфологической и функциональной перестройки в тяжёловодородной среде. С помощью, электронной микроскопии дейтерированных и обычных клеток микрококка (В. А. Ерёмин, Л. Н. Чекулаева, 1978) были выявлены существенные различия в морфологии полученных клеток (рисунок 6). Клетки, выращенные на Ни D-средах, имели в 2—3 раза более толстую клеточную стенку; чем конт рольные клетки, распределение в них нитей ДНК было неравномерным. На микрофотографии дейтерированных клеток хлореллы и микрококка видны как плотные (одинарная стрел ка), так и электронно-прозрачные участки (двойная стрелка), последние, вероятно до приготовления срезов, состояли из плотно упакованных мембран, наподобие мезосом. Кроме того, для дейтерированного микрококка (рисунок 6, в) было также характерно резкое изменение формы клеток и направления их деления. При этом видно образование сложных структур, со стоящих из 6—8 клеток. Наблюдавшееся деление не заканчивалось обычным расхождением дочерних клеток, а приводило к образованию атипичных клеток—«монстров», описанных Мозесом с соавт. (Moses et al., 1958).
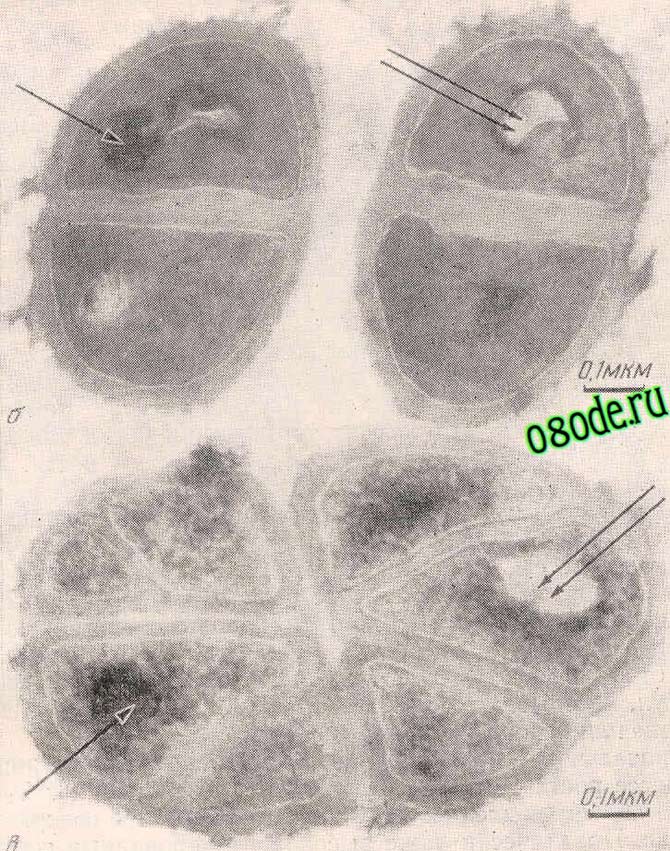
Рис. 6. Электронные микрофотографии клеток бактерий M. Lysodeikticus; б – клетки, выращенные на протонированной среде, в – дейтерированные клетки, выращенные на тяжёловодородной среде (по данным В. А. Ерёмина и Л. Н. Чекулаевой, 1978 ).
По-видимому, наблюдаемые морфологические изменения связаны с торможением роста дейтерированных клеток и обусловлены перестройкой в процессе адаптации к тяжёлой воде. Тот факт, что дейтерированные клетки микрококка кажутся более крупными (кажущийся размер в 2—4 раза превосходит размер протонированных клеток), является общебиологическим, так как наблюдается при выращивании целого ряда животных и растительных объектов (Денько; 1970).
Дейтерированные клетки адаптированных к максимальной концентрации тяжёлой воды в среде микроорганизмов – весьма удобные объекты для исследования. В процессе роста клеток на тяжёлой воде в них синтезируются макромолекулы, в которых атомы водорода в углеродном скелете почти полностью замещены на дейтерий. Такие дейтерированные макромолекулы претерпевают структурно-адаптационные модификации, необходимые для нормального функционирования клетки в тяжёлой воде.
Эти факты позволяют видеть некоторую аналогию между адаптацией к тяжёлой воде и адаптации к низким температурам. Ещё Юнг (Jung, 1967) на клетках Escherichia coli, помещенных в 98,6%-ную тяжёлую воду, показал, что эффект торможения роста тяжелой воды может быть компенсирован повышением температуры роста. Аналогия с охлаждением позволяет рассматривать адаптацию к тяжёлой воде, как адаптацию к неспецифическому фактору, действующему одновременно на функциональное состояние большого числа систем: превращение энергии, биосинтетические процессы, транспорт веществ, структуру и функции макромолекул. Возможно, что наиболее чувствительными к замене Н+ на D+ оказываются именно те системы, которые используют высокую подвижность протонов и высокую скорость разрыва протонных связей. Такими системами в клетке могут быть дыхательная цепь и аппарат биосинтеза макромолекул, которые располагаются в цитоплазматической мембране или находятся под ее контролем.
Аналогия между адаптацией к тяжёлой воде и температурной адаптацией очень важна для конструирования дейтерированных ферментов, которые смогут функционировать в условиях высоких температур. Такие стабильные дейтерированные ферменты необходимы в биотехнологии, медицине и сельском хозяйстве.
Это привело бы к ускорению обменных процессов в организме человека, а, следовательно, к увеличению его физической и интеллектуальной активности. Организм человека почти на 70% состоит из воды. И в этой воде 0,015% дейтерия. По количественному содержанию (в атомных процентах) он занимает 12-е место среди химических элементов, из которых состоит организм человека. В этом отношении его следует отнести к разряду микроэлементов. Содержание таких микроэлементов как медь, железо, цинк, молибден, марганец в нашем теле в десятки и сотни раз меньше, чем дейтерия. Что же случится, если удалить весь дейтерий? На этот вопрос науке еще предстоит ответить. Пока же несомненным является тот факт, что, меняя количественное содержание дейтерия в растительном или животном организме, мы можем ускорять или замедлять ход жизненных процессов.
Литературные источники:
Денько Е.И. Влияние тяжелой воды D2O на клетки животных, растений и микроорганизмы. Успехи совр. биол., 1970, т. 20. - № 1 (4). - с. 41 - 85.
Лобышев В.Н., Калиниченко Л.П. Изотопные эффекты D2O в биологических системах. М.: Наука, 1978. – 215 с.
Ерёмин В. А., Чекулаева Л. Н. Выращивание бактерий Micrococcus Lysodeikticus на дейтерированной среде, Микробиология, 1978
Crespi H. L. Biosynthesis and uses of per-deuterated proteins. in: Synt. and Appl. of Isot. Label. Compd. // Ed. R. R. Muccino. - Elsevier. - Amsterdam, 1986 - P. 111-112.
Katz J, Crespi H.L. // Pure Appl. Chem. - 1972. - V.32. - P. 221-250.
Daboll H. F., Crespi H. L., Katz J. J. // Biotechnology and Bioengineering. - 1962. - V. 4. - P. 281-297.
Jung K. // Studia Biophysica. - 1967 – V. 4. - 125
Мосин О. В., Карнаухова Е. Н., Пшеничникова А. Б., Складнев Д. А., Акимова О. Л. // Биотехнология. - 1993. - N 9. - С. 16-20.
Мосин О. В., Складнев Д. А., Егорова Т. А., Юркевич А. М., Швец В. И. // Биотехнология. - 1996. - N 3. - С. 3-12.
Мосин О. В., Складнев Д. А., Егорова Т. А., Юркевич А. М., Швец В. И. // Биотехнология. - 1996. - N 4. - С. 27-35.
Складнев Д. А., Мосин О. В., Егорова Т. А., Ерёмин С. В., Швец В. И. // Биотехнология. - 1996. - N 5. - С. 25-34.
Мосин О. В., Складнев Д. А., Егорова Т. А., Швец В. И. // Биоорганическая химия. - 1996. - Т. 22. - N 10-11. - С. 856-869.
Мосин О.В., Складнев Д.А., Швец. Исследование физиологической адаптации бактерий на тяжёловодородной среде. Биотехнология. 2000 г. N 8.
Бадьин В. И., Дробышевский Ю. В. и др. Отчет о НИР «Разработка препарата и способа его получения для стимуляции жизнедеятельности организма», Фирма «Мед-Чернобыль», 1993 г.
Бердышев Г.Д., Варнавский И.Н., Прилипенко В.Д. Аквабиотика – наука о роли воды в жизненных процессах. В кн.: Информоенергетика -го тисячол ття: соц олого-синергетичн та медично-еколог чн п дходи. Ки в – Кривий Р г: ЗАТ “ЗТНВФ “Коло”, 2003, с. 22 – 28.
Варнавский И.Н., Бердышев Г.Д., Прилипенко В.Д. Целебная реликтовая вода – открытие третьего тысячелетия. Вопросы химии и химической технологии, 2002, № 5, с. 168 – 174.