Тяжелая вода и адаптация к ней клеток
О. В. МОСИН
Московская государственная академия тонкой химической технологии им. М.В. Ломоносова, 117571, Москва, просп. Вернадского, д. 86.
Тяжёлая вода отличается от обычной воды молекулярной массой. В молекуле тяжёлой воды в отличие от обычной воды вместо двух атомов водорода, связанных ковалентной связью с атомом кислорода в молекуле эти два атома водорода замещены на дейтерий. Атом дейтерия, как изотоп водорода содержит одинаковое количество положительно заряженных частиц протонов, но различается от атома водорода содержанием нейтрально заряженных частиц нейтронов, которых в атоме дейтерия 2. Различие в нуклеарной массе атомов водорода и дейтерия и определяет те гидрофобные эффекты, которые тяжёлая вода оказывает на клетки и организм.
Важной проблемой является адаптация клетки к тяжёлой воде. Долгое время считалось, что тяжёлая вода несовместима с жизнью. Известно, что высокие концентрации тяжёлой воды в ростовой среде могут вызвать ингибирование жизненно-важных функций роста и развития многих микроорганизмов [1]. Несмотря на негативный биостатический эффект, оказываемый тяжёлой водой на клетки, некоторые бактерии устойчивы к высоким концентрациям тяжёлой воды в среде [2], в то время как растительные клетки могут нормально развиваться при концентрациях не более 50-75% тяжёлой воды [3], а клетки животных не более 35% тяжёлой воды [4].
Каков механизм клеточной адаптации к тяжёлой воде и могут ли клетки быть успешно адаптированы к росту и биосинтезу на средах с максимальными концентрациями тяжёлой воды? Для решения этих вопросов мы выбрали для наших экспериментов генетически маркированные штаммы-продуценты аминокислот, белков и нуклеозидов, относящиеся к различным таксономическим родам микроорганизмов [5-8]: факультативные метилотрофные бактерии Brevibacterium methylicum, облигатные метилотрофные бактерии Methylobacillus flagellatum, галофильные бактерии Halobacterium methylicum и бациллы Bacillus subtilis.
Результаты наших исследований изложены ниже:
Адаптация облигатных метилотрофных бактерий M. flagellatum к тяжёлой воде. Для проведения адаптации к тяжёлой воде были использованы представители двух различных таксономических групп метилотрофных бактерий, взятых из коллекции ГОСНИИ Генетики: лейцинпродуцирующий штамм облигатных метилотрофных бактерий Methylobacilus flagellatum, ассимилирующий метанол по 2-кето-3-дезокси-6-фосфоглюконатальдолазному варианту рибулозо-5-монофосфатного (РМФ) цикла фиксации углерода [9] и фенилаланинпродуцирующий штамм факультативных метилотрофных бактерий Brevibacterium methylicum, ассимилирующий метанол по РМФ-циклу фиксации углерода [10].
Для культивирования и адаптации данных штаммов метилотрофных бактерий использовали минимальные среды М9, содержащие в качестве источников дейтерия тяжёлую воду и [U-2H]метанол. В зависимости от физиологической потребности того или иного штамма бактерий в углеродном субстрате использовали 0.5-2% концентрации метанола в ростовых средах. Поскольку ба штамма ауксотрофы по лейцину и изолейцину для компенсации ауксотрофности эти аминокислоты добавляли в ростовые среды в концентрациях 10 мг/л. В обычных условиях культивирования на протонированных средах уровни накопления фенилаланина и лейцина в культуральных жидкостях штаммов-продуцентов достигали величины 0.8 и 1.0 г/л соответственно. 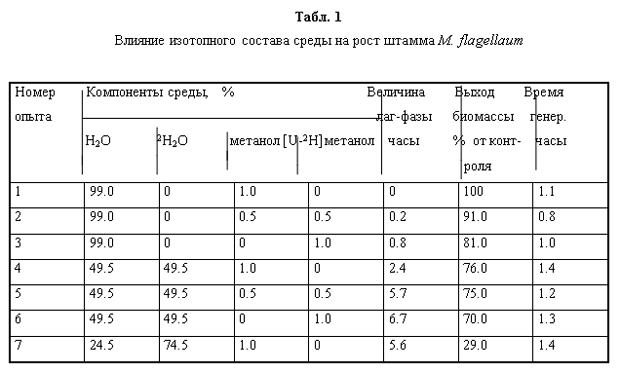
Для проведения адаптации был выбран ступенчатый режим увеличения концентрации тяжёлой воды в ростовых средах в присутствии 0.5-1% метанола/[U-2H]метанола , так как мы предположили, что постепенное привыкание организма к дейтерию будет оказывать благоприятный эффект на параметры роста и общее самочувствие клеток (табл. 1). При этом штамм M. flagellatum обнаружил повышенную чувствительность к тяжёлой воде: ингибирование скорости роста бактерий наблюдалось при концентрации тяжёлой воды в среде 74.5%, в то время как [U-2H]метанол не оказывал существенного влияния на скорость роста культуры. Так, на среде, содержащей 74.5% тяжёлую воду выход микробной биомассы составил 29%, что в 3.4 раза ниже, чем в контрольных экспериментах, когда использовали обычную воду и метанол (табл. 1, опыт 1), в то время как выход микробной биомассы на водной среде с 1% [U-2H]метанолом был снижен всего лишь в 1.2 раза. В связи с тем, что роста бактерий на более высокой концентрации тяжёлой воды достичь не удалось в дальнейших экспериментах использовали колонии клеток M. flagellatum, полученные со среды, содержащей 74.5% 2H2О.
Адаптация факультативных метилотрофных бактерий B. Methylicum к тяжёлой воде. Попытки адаптировать штамм B. methylicum к росту на тяжёлой воде при сохранении способности к биосинтезу фенилаланина привели к положительному результату. К данному штамму метилотрофных бактерий был применён специальный подход по адаптации, который заключался в серии из нескольких адаптационных пассажей исходной культуры на твёрдых агаризованных средах с 2% [U-2H]метанолом при ступенчатом увеличении концентрации тяжёлой воды в них (от 0 до 98% тяжёлой воды), как показано в табл. 2. При этом последовательно отбирали отдельные колонии, выросшие на средах, содержащих дейтерий. Затем их пересевали на среды с большей степенью дейтерированности, включая среду с 98% тяжёлой водой (степень выживаемости бактерий на максимальной тяжёловодородной среде составила не более 50%). За ходом адаптации следили по изменениям продолжительности лаг-фазы, времени клеточной генерации и выходов микробной биомассы, а также по максимальному уровню накопления фенилаланина в культуральной жидкости (рис. 1). 
Рис.1. Характеристики бактериального роста B. Methylicum (выход биомассы, время генерации и секреция фенилаланина) на средах со ступенчато увеличивающейся концентрацией тежёлой воды, соответствующим номерам экспериментов в таблице.
При росте клеток на обычной воде продолжительность лаг-фазы не превышала 20 ч, в то время как с увеличением концентрации тяжёлой воды в ростовых средах до 98% продолжительность лаг-фазы увеличивалась до 60 часов (таблица 2, опыт 10). Отмечено, что длительность времени клеточной генерации с увеличением концентрации тяжёлой воды в ростовых средах постепенно увеличивается, достигая 4,9 часов на среде с 98% тяжёлой воды и 2% [U-2H]метанолом (табл. 2, опыт 10). В отличие от тяжёлой воды, [U-2H]метанол не вызывал существенного ингибирования роста и не оказывал влияния на выходе микробной биомассы. Напротив, на максимально концентрированной тяжёлой воде водой выход микробной биомассы был снижен в 3.3 раза по сравнению с контролем. Важно то, что выход микробной биомассы и уровень накопления фенилаланина в культуральной жидкости при росте адаптированного к тяжёлой воде микроорганизма в максимально концентрированной тяжёлой воде изменяются по сравнению с контрольными условиями на 13 и 5%, т. е. незначительно (табл. 2, опыт 10’). 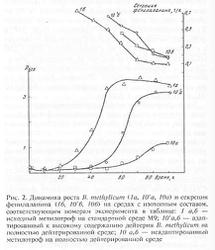
Рис.2. Динамики роста B. methylicum (1ф, 10’а, 10а) и секреция фенилаланина (1б, 10’б, 10б) на средах с обычной и тяжёлой водой: 1 а,б – исходный метилотроф на обычной воде; 10’а,б – адаптированный к тяжёлой воде микроорганизм; 10 а,б – неадаптированный метилотроф на среде с тяжёлой водой
Интересным фактом является и то, что адаптированные к тяжёлой воде клетки сохранили способность синтезировать и экзогенно продуцировать фенилаланин в ростовую среду. Причём общей особенностью биосинтеза фенилаланина в водно/тяжёловодородных средах было увеличение его продукции на раней фазе экспоненциального роста B. methylicum, когда выход микробной биомассы был незначителен (рис. 2). Во всех экспериментах наблюдалось ингибирование биосинтеза фенилаланина на поздней фазе экспоненциального роста и снижение его концентрации в ростовых средах. Согласно данным по микроскопическому исследованию растущей популяции микроорганизмов, подобный характер динамики секреции фенилаланина не коррелировал с качественными изменениями ростовых характеристик культуры на различных стадиях роста, что служило подтверждением морфологической однородности микробной популяции. Скорее всего, накопленный в процессе роста фенилаланин ингибировал ферменты собственного пути биосинтеза. Кроме того, не исключена возможность, что при ферментации без рН-статирования может происходить как обратное превращение секретируемого фенилаланина в интермедиаторные соединения его биосинтеза, так и ассимиляция фенилаланина клеткой для обеспечения собственных метаболических потребностей, что отмечено в других работах [11, 12].
Данные по исследованию культуральной жидкости методом ТСХ показали, что кроме фенилаланина штамм B. methylicum синтезирует и накапливает в культуральной жидкости (на уровне 5-6 ммоль) другие аминокислоты (аланин, валин, лейцин, изолейцин), присутствие которых также подтверждалось масс-спектрометрическим анализом метиловых эфиров их DNS-производных [13]. 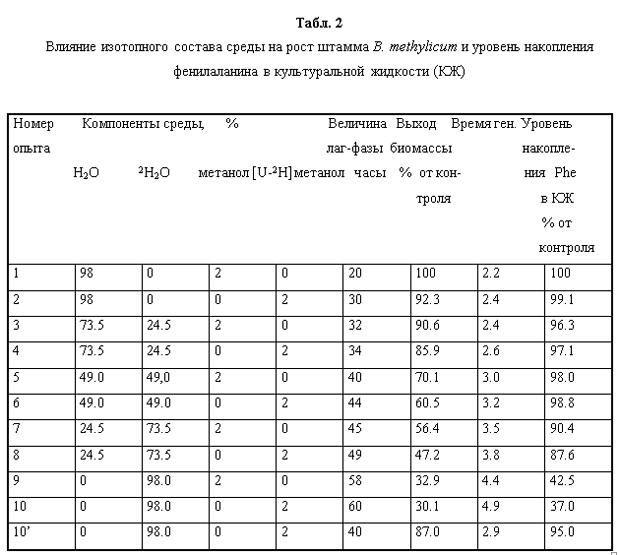
Так как данный штамм метилотрофных бактерий удалось адаптировать к максимальной концентрации тяжёлой воды, исследование принципиальной возможности использования гидролизатов его биомассы для культивирования других штаммов продуцентов представлялось весьма актуальным. Следует подчеркнуть, что усваиваемость биомассы метилотрофов клетками эукариот составляет 85-99%, а производительность метилотрофов, измеренная по уровню конверсии метилового спирта достигает 50% при этом учитывалось, что метилотрофные бактерии при росте на метаноле способны синтезировать большое количество полноценных белков (до 55% от веса сухого вещества), а также некоторое количество полисахаридов (до 10%), причем эта способность сохраняется при росте на средах, содержащих тяжёлую воду и [U-2H]метанол. Для выделения этих соединений из (2Н)меченой биомассы метилотрофных бактерий было необходимо проводить её гидролиз. Для этого использовали два метода гидролиза биомассы - щадящий гидролиз путём автоклавирования в 0.5 н. растворе дейтерохлорной кислоты (в тяжёлой воде) (30 мин, 08 ати) и исчерпывающий гидролиз биомассы в 6 н.дейтерохлорной кислоте (в тяжёлой воде) (24 часа, 110 0С). В предварительных экспериментах было показано, что по-первому варианту гидролиза биомассы реализуется гораздо большая питательность суспензии метилотрофных бактерий по сравнению с гидролизом в 6 н. дейтерохлорной кислоте. Поэтому мы отдали предпочтение этому методу проведения гидролиза биомассы.
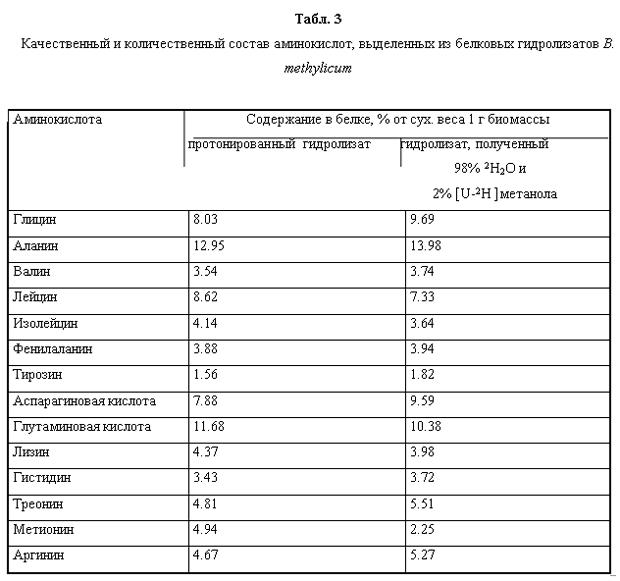
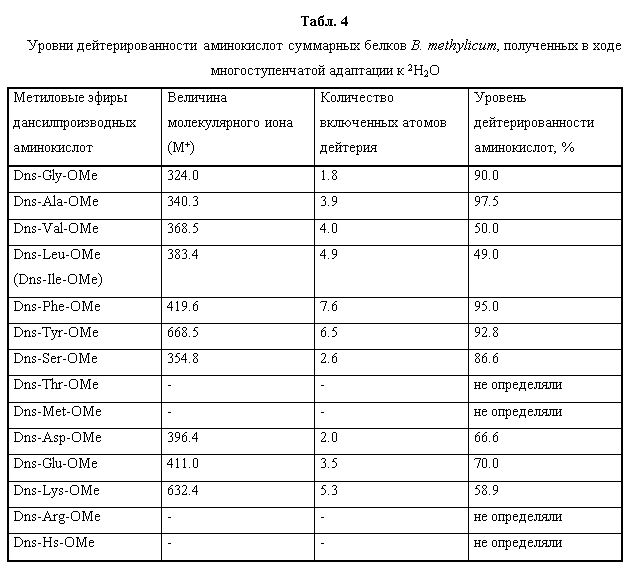
Вследствие того, что используемые в работе бактериальные штаммы были представлены их ауксотрофными по определенным аминокислотам формами, было необходимо оценить сколько данных аминокислот содержится в гидролизатах биомассы и каковы уровни их дейтерированности. В гидролизате биомассы, полученной с тяжёловодородной среды было зафиксировано небольшое снижение содержания лейцина, изолейцина, глутаминовой кислоты, лизина и метионина по сравнению с биомассой, полученной на обычной воде (табл. 3). Содержания аланина, аспарагиновой кислоты, треонина и аргинина в дейтерированном белке, напротив, немного превышают контрольные показатели, снятые в протонированной воде. Таким образом, достигнутый результат в опытах по адаптации B. methylicum к тяжёлой воде, позволил использовать гидролизаты его (2Н)меченой биомассы, полученной в ходе многоступенчатой адаптации к тяжёлой воде в качестве ростовых субстратов для выращивания бацилл Bacillus subtilis, а также штамма галофильных бактерий Halobacterium halobium. При этом, показателем, позволяющим надеяться на высокую эффективность включения дейтерия в продукты, синтезируемыми данными бактериальными штаммами, служит высокий уровень дейтерированности аминокислот суммарного белка этих бактерий, измеренный на метиловых эфирах DNS-производных аминокислот, за исключением лейцина и метаболически родственных с ним аминокислот, сниженные уровни дейтерированности которых объясняются эффектом ауксотрофности данного метилотрофного штамма в лейцине (табл. 4).
Адаптация бацилл B. subtilis к тяжёлой воде. В следующих опытах была исследована способность к росту на тяжёлой воде бациллярного штамма B. subtilis, продуцента инозина. Рост данного штамма лучше всего происходил на ГС 1 среде, содержащей в качестве источника углерода глюкозу, а в качестве источника ростовых факторов гидролизаты (2Н)меченой биомассы метилотрофных бактерий B. methylicum. 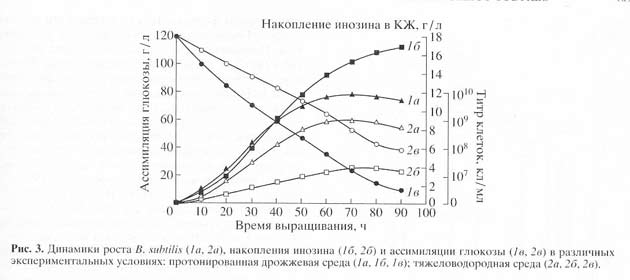
Рис.3. Динамики роста Bacillus subtilis (1а,2а), накопления инозина (1б,2Б) и ассимиляции глюкозы (1в,2в) на средах с обычной и тяжёлой водой: обычная вода (1а,1б,1в); тяжёлая вода (2а,2б,2в)
Данный штамм удалось адаптировать к тяжёлой воде путём рассева на твёрдую агаризованную среду ГС 1 с 99.9% тяжёлой воды. Он сразу обнаружил нормальный рост на тяжёлой воде. При культивировании адаптированного B. subtilis на жидкой ГС 1 среде, уровень накопления инозина в культуральной жидкости снижается по-сравнению с исходным штаммом. Например, при росте исходного штамма B. subtilis на среде, содержащей обычную воду и протонированную биомассу уровень накопления инозина в культуральной жидкости достигал величины 17 г/л после пяти суток культивирования (рис. 3). Вместе с тем уровень накопления инозина на ГС 1 среде, был снижен в 4.4 раза по сравнению с исходным штаммом на протонированной среде. Сниженный уровень продукции инозина на в этих условиях коррелирует со степенью конверсии глюкозы, которая на тяжёлой воде ассимилировалась не полностью, о чём свидетельствовали значительные количества накопленной в культуральной жидкости глюкозы после ферментации.
Поэтому было интересно оценить содержание глюкозы в гидролизатах биомассы B. subtilis. В состав гидролизатов внутриклеточных сахаров данного штамма входят глюкоза, фруктоза, рамноза, арабиноза, мальтоза и сахароза (табл. 5). Важно, что содержание глюкозы в образце, полученном с тяжёлой воды достигает 21.4%, т. е. значительно выше, чем для других сахаров. Содержания других сахаров в анализируемых образцах с тяжёлой водой существенно не отличаются от таковых для обычной воды, за исключением сахарозы, которая в дейтерированном образце не детектируется (табл. 5). 
Адаптация солелюбивых бактерий H. halobium к тяжёлой воде. В случае с H. halobium адаптацию проводили как на агаре, содержащим 99.9% тяжёлую воду с добавлением гидролизатов (2Н)меченой биомассы B. methylicum, путём рассева штамма до отдельных колоний, так и на жидкой ГС 2 среде. В обычных для этой культуры условиях культивирования (37 0С, на свету) в клетках синтезировался фиолетовый пигмент по всем характеристикам не отличающийся от нативного белка бактериородопсина.
Проведённые нами исследования свидетельствуют, что способность к адаптации к тяжёлой воде у разных родов и видов бактерий различная и может варьировать на примере метилотрофных бактерий в пределах даже одной таксономической группы. Из этого можно заключить, что адаптация к тяжёлой воде определяется как таксономической специфичностью микрооорганизмов, так и особенностями их метаболизма, функционированием различных путей ассимиляции субстратов, а также эволюционной нисшей, которую занимает исследуемый объект. При этом чем ниже уровень эволюционного развития организма, тем лучше он приспосабливается к присутствию дейтерия в среде. Так, из изученных объектов самыми примитивными в эволюционном плане являются галофильные бактерии, относящиеся к археобактериям, практически не нуждающие в адаптации к тяжёлой воде, чего нельзя сказать о метилотрофных бактериях, которые труднее адаптируются к тяжёлой воде. Для всех изученных микроорганизмов рост на тяжёлой воде сопровождался снижением ростовых характеристик а также уровня продукции секретируемых БАС.
Полученные для изученных микроорганизмов данные в целом подтверждают устойчивое представление о том, что адаптация к тяжёлой воде является фенотипическим явлением, поскольку адаптированные к тяжелой воде клетки возвращаются к нормальному росту и биосинтезу в протонированных средах после некоторого лаг-периода. По-видимому, метаболизм адаптированных клеток не претерпевает существенных изменений в тяжёлой воде. В то же время эффект обратимости роста на водно/тяжёловодородных средах теоретически не исключает возможности того, что этот признак стабильно сохраняется при росте в воде, но маскируется при переносе клеток на тяжёлую воду. Однако, здесь необходимо подчеркнуть, что для проведения адаптации играет немаловажную роль состав среды культивирования. При этом не исключено, что при проведении адаптации на минимальных средах, содержащих тяжёлую воду образуются формы бактерий, ауксотрофные по определенным ростовым факторам, например аминокислотам, и вследствие этого бактериальный рост ингибируется. Адаптация к тяжёлой воде происходит лучше всего именно на комплексных средах, содержащих широкий набор ростовых факторов и аминокислот, компенсирующих потребность бактерий в этих соединениях.
Можно предположить, что клетка реализует лабильные адаптивные механизмы, которые способствуют функциональной реорганизации работы жизненно-важных систем в тяжёлой воде. Так, например, нормальному биосинтезу и функционированию в тяжёлой воде таких биологически активных соединений, как нуклеиновые кислоты и белки способствует поддержание их структуры посредством формирования водородных (дейтериевых) связей в молекулах. Связи, сформированные атомами дейтерия различаются по прочности и энергии от аналогичных водородных связей. Различия в нуклеарной массе атома водорода и дейтерия косвенно могут служить причиной различий в синтезах нуклеиновых кислот, которые могут приводить в свою очередь к структурным различиям и, следовательно, к функциональным изменениям в клетке. Вероятнее всего, что ферментативные функции и структура синтезируемых белков также изменяются при росте клеток на тяжёлой воде, что может отразиться на процессах метаболизма и деления клетки. Некоторые исследователи сообщают, что после обратного изотопного (1Н-2H)-обмена ферменты не прекращают своей функции, но изменения в результате изотопного замещения за счет первичного и вторичного изотопных эффектов, а также действие тяжёлой воды как растворителя (большая структурированность и вязкость по сравнению с обычной водой) приводили к изменению скоростей и специфичности ферментативных реакций в тяжёлой воде [14-15].
Структурно-динамические свойства клеточной мембраны, которые в большинстве зависят от качественного и количественного состава липидов, также могут изменяться в присутствии тяжёлой воды. Так, сравнительный анализ липидного состава дейтерированных клеток B. subtilis, полученных при росте на тяжёлой воде показал различия в количественном составе мембранных липидов по сравнению с обычной водой (рис. 4). Примечательно, что в образце полученном с тяжёлой воды соединения, имеющие времена удерживания - 33.38; 33.74 и 33.2 мин не детектируются (рис. 4 б). Полученный результат, по видимому, объясняется тем, что клеточная мембрана является одной из первых органелл клетки, которая испытывает воздействие тяжёлой воды, и тем самым компенсирует реалогические параметры мембраны (вязкость, текучесть, структурированность) изменением количественного состава липидов.

Рис.4. Липидные профили бактерий B. subtilis, полученные на обычной (а) и тяжёлой воде (б): Анализ проводили на хроматографе Beckman Gold Systems (США); сорбент: Ultrasphere ODS 5 мкм; 4.6x250 мм; элюент: линейный градиент 5 мМ бисульфат калия-ацетонитрил; 100 об.% в течении 50 мин; скорость элюции: 0.5 мл/мин; детекция при длине волны 210 нм.
В общих чертах, при попадании клетки в дейтерированную среду из неё не только исчезает протонированная вода за счет реакции обмена вода-тяжёлая вода, но и происходит очень быстрый изотопный (1Н-2H)-обмен в гидроксильных, карбоксильных, сульфгидрильных и аминогруппах всех органических соединений, включая нуклеиновые кислоты, липиды, белки и сахара. Известно, что в этих условиях только С-Н связь не подвергается изотопному обмену и вследствие этого только соединения со связями типа С-2H могут синтезироваться de novo [16].
Кроме вышеобозначенных эффектов, возможное изменение соотношения основных метаболитов в процессе адаптации к тяжелой воде также может негативно сказываться на рост клетки.
Также не исключено, что эффекты, наблюдаемые при адаптации к тяжёлой воде связаны с образованием в тяжёлой воде конформаций молекул с иными структурно-динамическими свойствами, чем конформаций, образованных с участием водорода, и поэтому имеющих другую активность и биологические свойства. Так, по теории абсолютных скоростей разрыв С2H-связей может происходить быстрее, чем СH-связей, подвижность иона 2H+ меньше, чем подвижность Н+, константа ионизации тяжёлой воды несколько меньше константы ионизации обычной воды [17].
Суммируя полученные данные, можно сделать вывод, что чувствительности различных клеточных систем к тяжёлой воде отличны. С точки зрения физиологии, наиболее чувствительными к замене Н+ на 2H+ могут оказаться аппарат биосинтеза макромолекул и дыхательная цепь, т. е., именно те клеточные системы, которые используют высокую подвижность протонов и высокую скорость разрыва водородных связей.
Нам представляется выбор бактерий в качестве модельных объектов для данных исследований наиболее целесообразным, так как прокариоты как организмы, стоящие на более низких ступенях развития живого, наиболее лабильны в генетическом аспекте и тем самым быстрее реагируют и приспосабливаются к изменчивым факторам среды. В заключение следует подчеркнуть, что для того чтобы сделать более конкретные выводы о природе и механизме адаптации клеток к тяжелой воде, необходимы новые экспериментальные данные по физиологии и биохимии адаптированных клеток.
Материалы и методы:
Объектами исследования служили генетически маркированные штаммы-продуценты аминокислот, белков и нуклеозидов, полученные из Всероссийской коллекции промышленных микроорганизмов (ВКПМ) Государственного научно-исследовательского института генетики и селекции промышленных микроорганизмов:
1. Brevibacterium methylicum ВКПМ В 5652, лейцинзависимый штамм факультативных метилотрофных бактерий, продуцент фенилаланина.
2. Methylobacillus flagellatum КТ, изолейцинзависимый штамм облигатных метилотрофных бактерий, продуцент лейцина.
3. Bacillus subtilis В-3157, полиауксотрофный по гистидину, тирозину, аденину и урацилу штамм грамотрицательных бактерий, продуцент инозина.
4. Halobacterium halobium ЕТ 1001, пигментсодержащий штамм галофильных бактерий, способный синтезировать бактериородопсин.
Для приготовления сред и адаптации бактерий использовали тяжёлую воду (99.9% 2H), и дейтерохлористую кислоту (95.5% 2H) и [U2Н]метанол (97.5% 2H), полученные из Российского научно-исследовательского центра “Изотоп” (Санкт-Петербург, РФ). По необходимости тяжёлую воду очищали от вредных примесей, перегоняя её над перманганатом калия [18].
Стартовым материалом для культивирования галофильных бактерий и бацилл служила (2Н)меченая биомасса метилотрофных бактерий, полученная в условиях многостадийной адаптации на твердых агаризованных средах (2% агар) с 2% [U2Н]метанолом, содержащих ступенчато увеличивающиеся концентрации тяжёлой воды (от 0 до 98% тяжёлой воды). Полученную таким образом (2Н)меченую биомассу B. methylicum (выход составил 100 г по влажному весу с 1 л. среды) автоклавировали в 0.5 н. растворе дейтерохлорной кислоты (в тяжёлой воде) (08 ати, 30 мин), нейтрализовали 0.1 н. КОН (рН 7.0) и использовали далее в качестве источника ростовых факторов для адаптации и культивирования бацилл и галофильных бактерий.
При культивировании клеток микроорганизмов использовали следующие питательные среды (количества компонентов приведены в г/л):
1. Минимальная среда М9, на основе различных концентраций тяжёлой воды (см. табл. 1 и табл. 2) и добавками 0.5-2% метанола (в зависимости от физиологической потребности бактерий) или [U-2H]метанола: KH2PO4 3; Na2HPO4 6; NaCl 0.5; NH4Cl 1. Среду использовали для культивирования метилотрофных бактерий.
2. Гидролизная среда 1 (ГС1) для культивирования бацилл (на основе 99.9% тяжёлой воды): глюкоза 120; (2Н)меченая биомасса B. methylicum 25; NH4NO3 30; MgSO4 x 7H2O 20; мел 20.
3. Гидролизная среда 2 (ГС2) для культивирования галофильных бактерий (на основе 99.9% тяжёлой воды): NaCl 250; MgSO4 x 7H2O 20; KCl 2; CaCl2 x 6H2O 0.065; цитрат натрия 0.5; (2Н)меченая биомасса B. methylicum 20.
Культивирование метилотрофных бактерий и бацилл проводили при 370 С в колбах Эрленмейера вместимостью 250 мл с наполнением средой до 50 мл в условиях интенсивной аэрации по методикам [5-8], используя в качестве источников дейтерия тяжёлую воду и [U-2H]метанол. В случае с галофильными бактериями их культивирование проводили на тяжеловодородной среде при освещении лампами дневного света ЛБ-40. После 6-7 суток культивирования клетки отделяли центрифугированием (10000 об/мин, 20 мин). В культуральных жидкостях анализировали секретируемые аминокислоты и нуклеозиды.
Для выделения фракции суммарных белков биомассы клетки дважды промывали дистиллированной водой с последующим центрифугированием (10000 об/мин, 20 мин), экспонировали ультразвуком при 22 кГц (3 x 15 мин) и центрифугировали. Липиды и пигменты экстрагировали смесью органических растворителей хлороформ-метанол-ацетон (2:1:1) по методу Блайя и Дайера [19]. Полученный осадок высушивали до постоянного веса (10-12 мг) и использовали его в качестве фракции суммарных белков биомассы.
Суммарные белки биомассы гидролизовали в запаянных стеклянных ампулах в 50-ти кратном избытке 6 н. дейтерохлористой кислоты (в тяжёлой воде). Ампулы выдерживали при 1100 в течение 24 ч. После этого реакционную массу суспендировали в горячей дистиллированной воде, фильтровали. Гидролизат упаривали в роторном испарителе при 400 С. Остатки дейтеросоляной кислоты удаляли путем выдерживания в эксикаторе над твердым NaOH.
Для проведения гидролиза внутриклеточных полисахаридов 50 мг сухой делипидизованной биомассы помещали в круглодонную колбу вмесимостью 250 мл, добавляли 50 мл дистиллированной воды и 1.6 мл 25% дейтеросерной кислоты и кипятили с обратным водяным холодильником в течении 90 мин. По охлаждении реакционную смесь суспендировали в одном объёме горячей дистиллированной воды и нейтрализовали 2 н. раствором Ba(ОН)2 до рН 7.0. Выпавший осадок сульфата бария отделяли центрифугированием (15000 об/мин, 5 мин), супернатант декантировали и упаривали в роторном испарителе при 400 С.
Рост бактерий оценивали по способности к образованию отдельных колоний на поверхности твёрдых агаризованных сред, а также по величине оптической плотности суспензии клеток, измеренной на спектрофотометре Beckman-DU6 (США) при 540 нм в кварцевой кювете с длиной оптического пути 10 мм.
Тонкослойную хроматографию (ТСХ) проводили на пластинках Silufol UV-254 (Чехо-Словакия) или на пластинках с закреплённым слоем селикагеля фирмы Merck (Германия) в системах растворителей: изопропанол-аммиак, (7:3) (A) для аминокислот и н-бутанол-уксусная кислота-вода, (2:1:1), (Б) для инозина.
Определение содержания глюкозы в культуральной жидкости проводили глюкозооксидазным методом [20].
Секретируемые фенилаланин и инозин определяли на приборе Beckman DU-6 (США). Фенилаланин определяли при 540 нм в образцах культуральной жидкости, объёмом 10 мкл после обработки препаратов культуральной жидкости нингидрином, инозин - при 249 нм в образцах культуральной жидкости, объёмом 20 мкл.
Аминокислотный анализ гидролизатов суммарных белков биомассы проводили на приборе Biotronic LC 5001 (ФРГ); 230 x 3,2 мм; рабочее давление 50-60 атм; скорость подачи натрийцитратного буфера 18.5; нингидрина 9.25 мл/ч; детекция при 570 нм и 440 нм (для пролина).
Анализ углеводов осуществляли на жидкостном хроматографе Knauer (ФРГ), снабженным насосом Gilson (ФРГ) и рефрактометром Waters К 401 (ФРГ); неподвижная фаза: Separon NH2, 10 мкм; 10 x 250 мм; подвижная фаза: ацетонитрил-вода, (75:25); скорость подачи: 0.6 мл/мин.
Липиды анализировали на хроматографе Beckman Gold System (США), снабжённым насосом Model 166 (США) и детектором Model 126 (США); неподвижная фаза: Ultrasphere ODS 5 мкм; 4.6 x 250 мм; подвижная фаза: линейный градиент 5 мМ KH2PO4-ацетонитрил; 100% в течении 50 мин; скорость подачи: 0.5 мл/мин; детекция при 210 нм.
Уровни включения дейтерия в аминокислоты белковых гидролизатов определяли методом масс-спектрометрии электронного удара в виде метиловых эфиров N-диметиламинонафталин-5-сульфонильных (дансильных) производных аминокислот на приборе MB-80A (Hitachi, Япония) при ионизирующем напряжении 70 эВ.
Список литературы:
1. Crespi H. L. Biosynthesis and uses of per-deuterated proteins. in: Synt. and Appl. of Isot. Label. Compd. // Ed. R. R. Muccino. - Elsevier. - Amsterdam, 1986 - P. 111-112.
2. Katz J, Crespi H.L. // Pure Appl. Chem. - 1972. - V.32. - P. 221-250.
3. Daboll H. F., Crespi H. L., Katz J. J. // Biotechnology and Bioengineering. - 1962. - V. 4. - P. 281-297.
4. Crespy H. L. Stable Isotopes in the Life Sciences. - International atomic energy agency. - Vienna. - 1977. - P. 111-121.
5. Мосин О. В., Карнаухова Е. Н., Пшеничникова А. Б., Складнев Д. А., Акимова О. Л. // Биотехнология. - 1993. - N 9. - С. 16-20.
6. Мосин О. В., Складнев Д. А., Егорова Т. А., Юркевич А. М., Швец В. И. // Биотехнология. - 1996. - N 3. - С. 3-12.
7. Мосин О. В., Складнев Д. А., Егорова Т. А., Юркевич А. М., Швец В. И. // Биотехнология. - 1996. - N 4. - С. 27-35.
8. Складнев Д. А., Мосин О. В., Егорова Т. А., Ерёмин С. В., Швец В. И. // Биотехнология. - 1996. - N 5. - С. 25-34.
9. Kletsova L. V., Chibisova E. S., Tsygankov Y. D. // Arch. Microbiol. - 1988. - V. 149. - P. 441-446.
10. Nesvera J., Patek M., Hochmannova J., Chibisova E., Serebrijski I., Tsygankov Y., Netrusov A. // Appl. Microb. Biotechnol. - 1991. - V. 35. - P. 777-780.
11. Ворошилова Э. Б., Гусятинер М. М., Жданова Н. И. // Биотехнология. - 1989. - N 2. - С. 137-141.
12. Максимова Н. П., Олехнович И. Н. Регуляция биосинтеза ароматических аминокислот у метилотрофных бактерий: Биохимия и физиология метилотрофов. - Пущино, 1987. - С. 77-85.
13. Мосин О. В., Складнев Д. А., Егорова Т. А., Швец В. И. // Биоорганическая химия. - 1996. - Т. 22. - N 10-11. - С. 856-869.
14. Barksdale A. D., Rosenberg A. // Methods Biochem. Anal. - 1982. - V. 28. - P. 1-25.
15. Tuchsen E., Woodward C. K. // J. Mol. Biol. - 1985. - V. 185. - P. 421-430.
16. Perrin C. L., Arrhenius G. M. L. // J. Am. Chem. Soc. - 1982. - V. 104. - P. 6693-6699.
17. Covington A. K., Robinson R. A., Bates R. G. // J. Phys. Chem. - 1966. - V. 70. - P. 3820-3829
18. Кейл Б. Лабораторная техника органической химии. - М.: Мир, 1966. - С. 504.
19. Bligh E.G., Dyer W.J. // Can. J. Biochem. Physiol. - 1959. - V. 37. - N. 8. - P. 911-918.
20. Полюдек-Фабини Р., Бейрих Т. // Органический анализ. - Л. Химия. - 1981. - С. 515-516.