Изучение уровней включения изотопов 2Ни 13С в секретируемые аминокислоты метилотрофных бактерий
Эффективность использования дансильных и Z-производных аминокислот для масс-спектрометрических исследований была показана раннее [10, 16]. В данной работе уровни включения изотопов 2Н-и 13С в мультикомпонентные смеси аминокислот в составе культуральной жидкости и белковых гидролизатов определяли методом масс-спектрометрии электронного удара метиловых эфиров дансил-аминокислот или в виде Z-производных аминокислот после их препаративного разделения методом обращённо-фазовой ВЭЖХ.
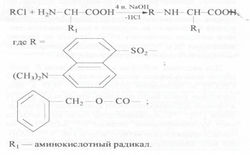
Для лизина, гистидина, тирозина, серина, треонина и цистеина наряду с монопроизводными было характерно образование ди-Z-(Dns)-производных: ди-Z,(Dns)-лизина, ди-Z,(Dns)-гистидина, О,N-ди-Z,(Dns)-тирозина, O,N-ди-Z,(Dns)-серина, O,N-ди-Z,(Dns)-треонина и N,S-ди-Z,(Dns)-цистеина (на схеме 1 эти произодные не показаны). Кроме этого, из аргинина синтезировался три-Z,(Dns)-аргинин.
Летучесть дансилпроизводных аминокислот при масс-спектрометрическом анализе повышали за счет дополнительной дериватизации по карбоксильной группе (этерификации) диазометаном. Выбор диазометана как этерифицирующего реагента был обусловлен необходимостью проведения реакции в мягких условиях, исключающих изотопный (1Н-2Н) обмен в ароматических аминокислотах.
Данные масс-спектрометрии по уровням включения стабильных изотопов 2Н-и13С в бензилоксикарбонильные производные аминокислот в пределах одинаковых концентраций тяжёлой воды в среде не отличались от таковых, полученных для метиловых эфиров дансиламинокислот (точность определения уровней изотопного включения в аминокислоты данным методом составляет +5). Данные по уровням включения 2Н-и 13С секретируемые аминокислоты исследуемых штаммов приведены в таблице 3. Для введения дейтерия в аминокислоты B. methylicum использовали минимальные среды с 2 об.% СН3ОН и различным содержанием 2Н2О в них, поскольку раннее было показано, что вклад С2Н3О2Н в уровни дейтерированности аминокислот незначителен [10].
13С-аминокислоты были получены за счёт культивирования штамма M. flagellatum на среде, содержащей обычную воду и 1 об.% к 13CH3OH. Во всех анализируемых образцах культуральной жидкости не зависимо от рода штамма присутствовали аланин, валин, лейцин (изолейцин) и фенилаланин (см. табл. 3). В культуральной жидкости M. flagellatum в дополнение вышеназванным аминокислотам также накапливался глицин.
Таблица 3.
Уровни включения 2Н и 13С в секретируемые аминокислоты B. methylicum и M. flagellatum (данные получены для Z-,и Dns-производных аминокислот)*.
- Аминокислоты; Содержание 2Н2О в среде, об% 24,5 49,0 73,5 98,0; 1 % 13СН3ОН
- Глицин; -; -; -; -; 60,0
- Аланин; 24,0; 37,5; 62,5; 77,5; 35,0
- Валин; 20,0; 46,3; 43,8; 58,8; 50,0
- Лейцин (изолейцин); 15,0; 47,0; 46,0; 51,0; 38,0
- фенилаланин; 15,0; 27,5; 51,3; 75,0; 95,0
* Данные по включению 2Н в аминокислоты приведены для B. methylicum при росте на средах, содержащих 2 об.% CH3OH и 24,5; 49,5; 73,5; 98,0 об.% 2Н2О.
Данные по включению 13С приведены для M. flagellatum при росте на среде, содержащей 1 об.% 13СН3ОН и 99 об.% Н2О.
Во всех опытах наблюдалось специфичное возрастание уровней изотопного включения дейтерия в индивидуальные аминокислоты культуральной жидкости при ступенчатом увеличении концентраций тяжёлой воды в ростовой среде (табл. 3). Как видно из таблицы 3, для аминокислот из культуральной жидкости, количество включенных атомов дейтерия по углеродному скелету молекулы варьирует в пределах 49 об.%-ной концентрации 2Н2O и составляет для фенилаланина 27,5%, аланина - 37,5%, валина - 46,3%, лейцина (изолейцина) - 47%.
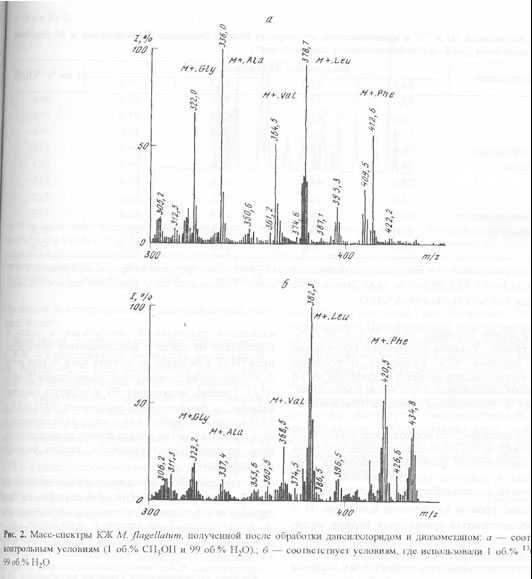
Как и следовало ожидать, для получения 13С-аминокислот за счет микробной биоконверсии 13СН3ОН, предварительная адаптация не является необходимым этапом, поскольку этот изотопный субстрат не оказывает существенного влияния на ростовые и биосинтетические характеристики метилотрофов. Масс-спектр культуральной жидкости M. flagellatum, полученной после обработки дансилхлоридом и диазометаном со среды, содержащей 1 об.% 13СН3ОН и 99 об.% Н2О показан на рис. 2,б (Масс-спектр приведён относительно контрольных условий, где использовали обычную воду и метанол (а)). Как видно из рис. 2,б, в дериватизованной культуральной жидкости М. lagellatum детектируются обогащённые изотопом 13С пики молекулярных ионов производных аминокислот с М+. при m/z 337,4; 368,5; 382,3; 420,5, которые соответствуют по массе аланину, валину, лейцину (изолейцину) и фенилаланину. Так как отношения массы к заряду m/z для лейцина (изолейцина) в масс-спектрах электронного удара метиловых эфиров дансиламинокислот совпадают, то вследствие этого нельзя точно идентифицировать структуру соединения данным методом. Кроме вышеназванных пиков молекулярных ионов, в масс - спектре фиксируется пик с М+. при m/z 323,2 (вместо m/z 322,0 в контроле), соответствующий метиловому эфиру дансил-глицина.
В связи с тем, что штамм B. methylicum был ауксотрофом по лейцину, а другой штамм M. flagellatum - ауксотрофом по изолейцину, было интересно изучить как изменяются уровни включения дейтерия в этих аминокислотах. Для этого лейцин добавляли в ростовую среду B. methylicum, приготовленную на основе 98 об.% 2Н2О в немеченном виде. В случае с M. flagellatum изолейцин добавляли в среду, приготовленную из обычной воды и 1 об.% 13СН3ОН. Как показали наши исследования, в условиях ауксотрофности по лейцину (изолейцину) уровень изотопного обогащения лейцина (изолейцина), а также метаболически связанных с ними аминокислот ниже, чем для других аминокислот. Так, при росте B. methylicum на среде, содержащей 98 об.% 2Н2О и немеченный L-лейцин, уровни включения дейтерия в лейцин (изолейцин) составили 51,0%, аланин - 77,5%, валин - 58,8% (табл. 3). Уровень включения дейтерия в фенилаланин в этих условиях составил 75%. Эта же особенность проявляется при росте M. flagellatum на среде с 1 об.% 13СН3ОН и добавкой немеченного L-изолейцина. Как видно из таблицы 3, в отличие от фенилаланина (уровень изотопного обогащения - 95%), уровни включения изотопа 13С в лейцин (изолейцин), аланин и валин составили 38,0; 35,0; 50,0 % соответственно. Уровень изотопного обогащения глицина составил 60%. Суммируя полученные данные, можно сделать вывод о сохранении минорных путей метаболизма, связанных с биосинтезом лейцина (изолейцина) de novo.