Использование ауксотрофных мутантов бактерий для получения изотопномеченных аминокислот и белков
Использование ауксотрофных по определённым аминокислотам форм микроорганизмов для биосинтеза изотопномеченых аминокислот и белков стало настолько популярным в биотехнологии, что сегодня его следует рассматривать как отдельное направление. Селективность мечения белков достигается в результате добавления в ростовую среду меченого аналога соответствующей аминокислоты или её предшественника, по которым штамм ауксотрофен и которые, вследствие этого, непосредственно или через de novo биосинтетический цикл предшественников заменяют в белке нативную аминокислоту. Аналогичный принцип может применяться и при получении изотопномеченых аналогов аминокислот. При этом ауксотрофные штаммы могут относиться к различным таксономическим группам микроорганизмов, включая метаногенные и метилотрофные бактерии, биотехнологический потенциал которых для получения изотопномеченых аминокислот в настоящее время общепризнан. Так, метаногенные бактерии, относящиеся к группе облигатных анаэробов, которые получают энергию за счет ассимиляции газовой смеси (H2-CO2) [88, 89], чаще всего используют для получения [13C]аминокислот. Причём эффективности мечения аминокислот 13С добиваются за счёт получения и использования ацетатзависимых мутантов метаногенных бактерий, неспособных синтезировать ацетил-СоА из СО2 и вследствие этого для роста которых необходим экзогенный ацетат [90].
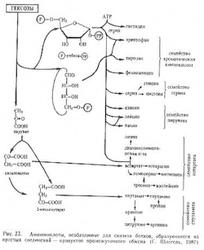
[13C] Аланин. Включение углерода 13С в молекулу аланина происходило за счет реакции карбоксилирования ацетил-СоА до пирувата. Такой путь биосинтеза был продемонстрирован для других таксономических родов и видов метаногенных бактерий [95].
[13C] Серин и [13C] глицин. Характер распределения изотопа 13C в молекулах серина и глицина был объяснён частичным фосфорилированинем пирувата до фосфопирувата и образованием 3-фосфоенолпирувата по гликогенному пути ассимиляции углерода. Подтверждением этому служат значительные уровни активности ферментов - фосфоенолпируватсинтетазы, енолазы и 2-фосфоглицератмутазы, которые были обнаружены в клеточных экстрактах других метаногенов, например, Methanobacterium thermoautotrophicum [96].
[13C] Аспарагиновая кислота, [13C] треонин и [13C] метионин. Присоединение 13C-метки по атому углерода -карбоксильной группы аспартата, происходящего из C1-ацетата и по -углеродному атому С2-ацетата и включение изотопа 13С в карбоксильные группы аминокислот из СО2, свидетельствовало о том, что биосинтез аспартата в этой бактерии происходил через цикл трикарбоновых кислот в результате ферментативного карбоксилирования пирувата до оксалоацетата. Распределение метки в треонине и метионине происходило в соответствии с путем биосинтеза этих аминокислот из аспартата. Атом углерода в метильной группе молекулы метионина происходил из СО2.
[13C] Лизин. Распределение метки 13С в молекуле лизина свидетельствовало о том, что лизин синтезировался из пирувата и аспартата по типичному для бактерий диаминопимелиновому пути [97].
[13C]Глутаминовая кислота, [13C]аргинин и [13C]пролин. В молекуле глутаминовой кислоты изотопная метка детектировалась в С и C положениях углеродного скелета молекулы. Атомы углерода при карбоксильной СООНгруппе молекулы глутаминовой кислоты и в -положении происходили из СО2. Этот результат свидетельствовал о том, что цикл трикарбоновых кислот приводил к образованию -кетоглутарата. Распределение 13C-метки в молекулах аргинина и пролина аналогично таковому в глутаминовой кислоте.
[13C] Лейцин, [13C] валин и [13C] изолейцин. Характер изотопного включения 13С в молекулы лейцина и валина свидетельствовал об их образовании из -ацетолактата, в то время как биосинтез изолейцина отличался от ожидаемого пути биосинтеза этой аминокислоты из треонина. В клетках M. hungatei изолейцин образовывался из ацетата. Аналогичный путь биосинтеза изолейцина был обнаружен у спирохеты [98], у лейцинассимилирующего мутанта Serratia marcescens [99], и у мутанта Saccharomyces cerevisiae, у которого дефектен ген треониндезаминазы [100].
[13C] Фенилаланин и [13C] тирозин. Меченые позиции углерода в молекулах фенилаланина и тирозина полностью совпадали с типичным для бактерий путем биосинтеза этих аминокислот из шикимовой и хоризмовой кислот [101].
[13C] Гистидин. Атом углерода в положении C имидазольного кольца гистидина происходил из СО2. Углеродный атом в положении С имидазольного кольца гистидина был замещён на изотоп 13С с участием С2ацетата.
Табл. 1
Параметры роста некоторых используемых в биотехнологии штаммов метилотрофных бактерий
- Штаммы бактерий; Молярный выход биомассы, г/моль метанола; Удельная скорость роста, ч-1; Эффективность конверсии углерода метанола, %; Количество потребленного азота, %
- Рибулозо-5-монофосфатный путь ассимиляции углерода
- Pseudomonas C1; 17,3; 0,49; 67,5; 13,2
- Pseudomonas methanolica; 17,0; 0,63; 66,5; 11,0
- Methylomonas methanolica; 15,7; 0,52; 62,0; 11,7
- Сериновый путь ассимиляции углерода
- Pseudomonas 1; 12,1; 0,176; 47,5; 11,37
- Pseudomonas 135; 12,1; 0,14; 47,5; 11,37
- Pseudomonas AM1; 9,8; 0,093; 37,6; 11,20
- Pseudomonas M27; 13,1; 0,108; 51,0; 9,40
- Pseudomonas roseus; 13,1; 0,15; 51,0; 10,60
Другими перспективными источниками изотопномеченых аминокислот и белков признаны метилотрофные микроорганизмы, которые в таксономическом аспекте представлены грамположительными, грамотрицательными бактериями и дрожжами, интерес к которым в настоящее время все возрастает благодаря разработке новых технологий химического синтеза метанола [102]. Эти бактерии привлекают внимание исследователей прежде всего как дешевые источники микробного белка и аминокислот [103, 104]. Знание путей бактериального метаболизма позволяет осуществлять направленное введение изотопной метки в молекулы аминокислот. Как известно, метилотрофные бактерии окисляют метанол с использованием фермента - метанолдегидрогеназы, последующие окислительные реакции катализируют формальдегиди формиатдегидрогеназа [105-108]. Лишь затем продукт окисления метанола в виде формальдегида фиксируется клеткой одним из двух путей ассимиляции углерода: рибулозо-5-монофосфатным и сериновым [109, 110]. Основные параметры роста некоторых используемых в биотехнологии штаммов метилотрофных бактерий представлены в таблице 1.
Ауксотрофные штаммы метилотрофных бактерий начали эффективно использовать для получения [2H]и [13C]аминокислот. Для этих целей перспективно осуществлять биологическую конверсию дешёвых низкомолекулярных меченых субстратов - (13С)метанола, (2Н)метанола и 2Н2O в дорогостоящие меченые БАС в клетках метилотрофов [111-113]. Традиционным подходом при этом остаётся выращивание соответствующих штаммов-продуцентов аминокислот, устойчивых к росту на средах, содержащих стабильные изотопы водорода, углерода, азота и др. В работах [114, 115] сообщается о получении [13C]аминокислот (уровни включения стабильных изотопов в молекулах варьируют от 30% для L-[13C]лейцина до 90% для L-[13C]фенилаланина) за счёт использования ауксотрофных по L-изолейцину бактерий Methylobacillus flagellatum.
Заслуживает упоминания то, что (13С)метанол в отличие от 2Н2О не оказывает существенного биостатического эффекта на ростовые и биосинтетические характеристики метилотрофов [116], поэтому данный подход можно эффективно использовать для введения в молекулы синтезируемых БАС двойной изотопной метки (например, введение изотопа 13С в молекулы на фоне максимальных концентраций 2Н2О в ростовых средах). В работе [117] были получены [2H]и [13С]аминокислоты с разными уровнями изотопной обогащённости при росте ауксотрофного по L-лейцину штамма факультативных метилотрофных бактерий Brevibacterium methylicum и ауксотрофного по L-изолейцину штамма облигатных метилотрофных бактерий Methylobacillus flagellatum на минимальных средах с (13С)метанолом, (2Н)метанолом и 2Н2О. [13C]и [2Н]аминокислоты разного уровня изотопной замещённости выделяли как из культуральных жидкостей, полученных после выращивания бактерий на средах с соответствующими изотопномечеными субстратами, так из гидролизатов белков биомассы. Биосинтетически полученные [2H]и [13С]аминокислоты представляли собой смеси, в которых присутствовали изотопнозамещённые формы молекул, различающиеся количеством атомов водорода и углерода, замещённых на 2Н и 13С. При этом распределение зависело как от общего включения изотопа в молекулу, так и от способа их получения. Данные по суммарным уровням включения стабильных изотопов в молекулы секретируемых аминокислот и аминокислотные остатки суммарных белков биомассы B. methylicum и M. flagellatum представлены в таблице 2. Эти исследования показали, что в условиях ауксотрофности по лейцину уровень изотопного обогащения лейцина, а также метаболически связанных с ним аминокислот немного ниже, чем для других аминокислот, вероятно, за счёт сохранения минорных путей метаболизма, связанных с биосинтезом данных аминокислот de novo. Так, при выращивании B. methylicum на среде, содержащей 98% 2Н2О и немеченый L-лейцин, уровни включения дейтерия в индивидуальные аминокислоты культуральной жидкости составили 51% для лейцина/изолейцина, 58,8% для валина, в то время как уровни изотопного включения для аланина составили 77,5%, для фенилаланина -75% (табл. 2).
Табл. 2
Суммарные уровни включения стабильных изотопов в молекулы секретируемых аминокислот и аминокислотные остатки суммарных белков биомассы B. methylicum* и M. flagellatum**.
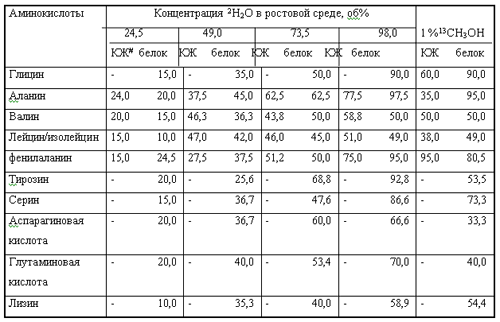
*Данные по включению дейтерия в молекулы аминокислот приведены для B. methylicum при росте на средах, содержащих 2% CH3OH и 24,5; 49,0; 73,5; 98,0% 2Н2О
**Данные по включению 13С приведены для M. flagellatum при росте на среде, содержащей обычную воду и 1% 13СН3ОН.
#Термином КЖ обозначены культуральные жидкости, полученные после отделения клеток из ростовых сред
Аналогичная корреляция наблюдалась и в аминокислотах белковых гидролизатов. Уровни включения 2Н и 13С в метаболически связанных аминокислотах в пределах одинаковых концентраций меченых субстратов, обнаружили определённую коррелляцию: уровни изотопного включения для валина и лейцина (семейство пирувата), фенилаланина и тирозина (семейство ароматических аминокислот) коррелировали (табл. 2). Уровни изотопного включения для глицина и серина (семейство серина), аспарагиновой кислоты и лизина (семейство аспарагина) также имели близкие величины. Важным результатом являются высокие уровни включения стабильных изотопов 2Н и 13С в молекулы полученных аминокислот. В настоящее время исследования по изучению биотехнологического потенциала метилотрофных бактерий для направленного синтеза изотопномеченых аминокислот и других БАС продолжаются.