Вода сохраняет приобретенную структуру несколько часов, правда, теряет ее при нагревании. Но наибольшей биологической силой обладает только что растаявшая вода, когда в посуде еще плавают отдельные льдинки.
Заморозить сырую воду в обычном холодильнике - налить кастрюлю или миску и поставить на лист фанеры или картона в морозилку. После замерзания вытащить и оставить таять при комнатной температуре.
Когда вода начинает замерзать, удалите только что образовавшуюся корочку льда. Это дейтерий, он замерзает в первую очередь.
После того как заледенеет основная маса воды, ополосните застывший кусок под краном струёй холодной воды. Кусок должен стать прозрачным, так как вода удалит изо льда вредные примеси. Далее растопите лёд и пейте полученную талую воду.
Небольшое количество жидкости нагревают до температуры 94 – 96 0 С. Вода ещё не кипит, но уже струйками поднимаются пузыри. В этот момент кастрюлю снимают и резко охлаждают, затем замораживают и оттаивают воду. Приготовленная таким способом талая вода проходит все фазы своего круговорота в природе: испаряется, охлаждается, замерзает и тает. Эта вода особенно полезна – она обладает огромной внутренней энергией.
Издавна считалось, что употребление талой воды способствует омоложению организма. Талая вода отличается от обычной своей структурой, более сходной со структурой протоплазмы наших клеток. Свойства талой воды сохраняются до 12 часов. Получить талую воду можно, замораживая обычную водопроводную воду в морозильной камере холодильника.
Для профилактики и лечения сосудистых расстройств применяют по 2-3 стакана холодной талой воды (можно с кусочками льда). Первый стакан выпивают рано утром за час до еды, остальные - в течение дня, за час до очередного приема пищи. Минимальная доза, оказывающая эффект, - 4-6 г талой воды на 1 кг веса. В ряде случаев дозу следует увеличить (если болезнь запущена, при ожирении, нарушении обмена веществ).
Из книги Соловьевой Веры Андреевны "Золотой ус: целительные рецепты"
Протиевая вода. Она готовится так же, как и талая, но при этом имеются существенные отличия. Ввиду того, что вода содержит несколько изомеров, желательно избавиться от неблагоприятных и отобрать те, на которых наш организм лучше всего "работает".
Методика отбора основана на том, что изомеры тяжелой воды замерзают первыми при температуре +3,8 °С, а наиболее благоприятные для организма - при 0 °С. Поэтому первый ледок, образующийся при замораживании воды, содержит в основном тяжелые изомеры (дейтерий) и его необходимо выбрасывать.
При дальнейшем замораживании вода, превращаясь в лед, вытесняет всю растворенную в ней грязь в незамерзшую часть. Там же собираются и легкие изомеры, которые замерзают при более низких температурах. Естественно, ни легкие изомеры воды, ни отжатая в нее грязь не нужны организму. Их необходимо выбрасывать.
Какой метод получения талой воды применять вам, решайте сами, дорогие читатели. Ниже приводятся полезные советы и рекомендации как правильно приготовить и применять талую воду.
Для приготовления талой воды не следует брать природный лед или же снег, поскольку они, как правило, загрязнены и содержат много вредных веществ.
Для замораживания воды лучше использовать пластмассовые банки, предназначенные для хранения питьевой воды. Стеклянные емкости могут разорваться, так как при замерзании вода расширяется и увеличивается в объеме.
Не следует замораживать воду в металлическом сосуде, поскольку это значительно снижает эффективность ее действия.
Ни в коем случае нельзя получать талую воду растоплением снеговой шубы на морозильнике, т.к. этот лёд может содержать вредные вещества и хладагенты и, кроме того, иметь неприятный запах.
Техника получения талой воды заключается в различных скоростях замерзания чистой воды и воды, содержащей примеси. Экспериментальным путём установлено, что медленно застывая, лед интенсивно захватывает примеси в начале и в конце замерзания. Поэтому при получении льда нужно отбросить первые образовавшиеся льдинки, а затем, после замерзания основной части воды, слить незамерзшие остатки.
Свежую талую воду можно получать в домашних условиях. Но для этого необходимо придерживаться некоторых общих правил.
Талая вода приготавливается из предварительно очищенной питьевой воды, которая заливается в чистые, плоские сосуды на 85% их объема.
Посуда для приготовления талой воды плотно закрывается и помещается в морозильные камеры до полного замерзания.
Эффект Мпембы (Парадокс Мпембы) — парадокс, который гласит, что горячая вода при некоторых условиях замерзает быстрее, чем холодная, хотя при этом она должна пройти температуру холодной воды в процессе замерзания. Данный парадокс является экспериментальным фактом, противоречащим обычным представлениям, согласно которым при одних и тех же условиях более нагретому телу для охлаждения до некоторой температуры требуется больше времени, чем менее нагретому телу для охлаждения до той же температуры.
Этот феномен замечали в своё время Аристотель, Френсис Бэкон и Рене Декарт, однако лишь в 1963 году танзанийский школьник Эрасто Мпемба установил, что горячая смесь мороженого замерзает быстрее, чем холодная.
Будучи учеником Магамбинской средней школы в Танзании Эрасто Мпемба делал практическую работу по поварскому делу. Ему нужно было изготовить самодельное мороженое - вскипятить молоко, растворить в нем сахар, охладить его до комнатной температуры, а затем поставить в холодильник для замерзания. По-видимому, Мпемба не был особо усердным учеником и промедлил с выполнением первой части задания. Опасаясь, что не успеет к концу урока, он поставил в холодильник еще горячее молоко. К его удивлению, оно замерзло даже раньше, чем молоко его товарищей, приготовленное по заданной технологии.
В целительской и бытовой практике большую популярность имеет, так называемая, талая вода. Под талой водой понимается вода, полученная из растаявшего снега или льда. При этом считается, что талая вода благотворно влияет на живые организмы и растения, оказывая на них некоторое стимулирующее воздействие. При этом обычно делается акцент на воде, полученной в период весеннего естественного снеготаяния. Именно в этот период были замечены ее более активные, по сравнению с обычной водой, воздействия на живые организмы и растения.
Вода имеет очень большое значение в жизни растений, животных и человека. Происхождение жизни на Земле обязано воде. В организме вода представляет собой среду, в которой протекают химические процессы, обеспечивающие жизнедеятельность организма; кроме того, вода принимает участие в целом ряде биохимических реакций как растворитель.
Вода очень необычная по своим физико-химическим свойствам субстанция. Плотность воды при переходе ее из твердого состояния в жидкое не уменьшается, как у других веществ, а возрастает. При нагревании воды от 0 до 4 °С плотность ее также увеличивается. При 4 °С вода имеет максимальную плотность, при дальнейшем нагревании ее плотность уменьшается. Это свойство воды очень ценно для жизни.
Кто-нибудь делал легкую воду методом В.А. Каргополова? Я нашла его патент на изобретение. Но не понятно, как сделать.
________
К сожалению, патента на получение легкой воды В.А. Каргополова я не нашел в патентном поиске. В других источниках содержится информация околонаучного содержания. Так что, если у вас имеется какая-либо информация, описание устройства и чертежи, вы можете выложить их на сайте. Изобретение (если таковое имеется) относится к усовершенствованию метода получения воды замораживанием-оттаиванием путем монтирования специальной т.н. дренажно-эвакуационной трубки, через которую удаляются примеси. Главная проблема, сопровождающая процесс приготовления легкой воды – это примеси в обычной воде тяжелой воды D2O.
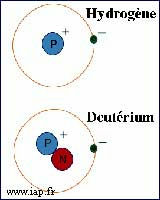
Тяжёлая вода по химическому составу представляет собой оксид дейтерия D2O с кислородом природного изотопного состава 16О. Строение молекул D2O такое же, как молекул Н2O, с очень малым различием в значениях длин ковалентных связей и углов между ними.
в одном из ваших ответов употребляющих лёгкую воду, к.х.н. О.В. Мосиным было сказано, что ".....У нас в России лёгкую воду производит одна единственная фирма “Лангвей”...." Мои поиски не увенчались успехом я нашла производителя OOO МТК Айсберг. Можно ли уточнить кто производит в нашей стране легкую воду.. Очень не хочется нарваться на подделку так как цена на эту воду достаточно высокая..
Страничка посвящена влиянию воды, освобожденной от тяжелых изотопов, на здоровье человека. Конкретнее - доступному способу получения такой воды и опыту ее использования, личному и того круга людей, которые приняли участие в "эксперименте".
Слово "изотоп" у большинства людей наверняка ассоциируется с чем-нибудь опасным или, по крайней мере, неприятным. Радиоактивное заражение, радиология... Но, заглянув в таблицу элементов, неприятные ассоциации легко перебороть: кислород, азот (воздух), хлор (поваренная соль), да и практически все элементы в окружающих нас веществах - смеси изотопов. И ничего страшного.
Действительно, более тяжелый изотоп - это один или два лишних нейтрона в ядре. Что за дело до них биологическим объектам, каковыми мы являемся? Хотя... Основа биологии - химия, все-таки. Подвижность молекулы, да и другие важные в обмене веществ свойства, зависят от массы входящих в нее атомов. А у такого важного и распространенного элемента как водород второй по распространенности изотоп вдвое тяжелее основного.
Легкая вода с сохраненным солевым составом природных источников и здоровье человека. Вода является одной из основ жизни на Земле и присутствует во всех биологических и абсолютном большинстве природных объектов. Её природные функции чрезвычайно многообразны, свойства и качество воды отражаются на состоянии всех живых существ.
Вода в природе растворяет минералы, химические вещества, соли, и содержит взвесь биологических и неорганических объектов. При превышении некоторых концентраций растворенных веществ и взвесей, они становятся загрязнителями воды, применяемой в жизни человека. В современных условиях питьевая вода может содержать различные загрязнения, общее количество которых постоянно возрастает, и насчитывается до 30000. Часть этих загрязнений образуют живые организмы; одноклеточные водоросли, микробы, вирусы или продукты распада живой материи как хлорофилл и аминокислоты. Кроме того, в воде могут присутствовать вещества, абсолютно чужеродные большинству живых существ - ксенобиотики.
Лёгкая вода – это природная вода, очищенная от тяжёлой воды.
Изотоп водорода, дейтерий, отличающийся наличием в ядре «лишнего» нейтрона, может образовывать с кислородом молекулу воды. Такая вода, в молекуле которой атом водорода замещён атомом дейтерия, называется тяжёлой. Содержание дейтерия в различных природных водах очень неравномерно. Оно может меняться от 90 ppm (частей на миллион частей основного раствора) – это вода из Антарктического льда, - самая лёгкая природная вода, до 180 ppm в воде из подземных газовых пластов и закрытых водоёмов Сахары. Лёгкая вода является побочным продуктом производства тяжёлой воды, используемой в атомной промышленности в качестве замедлителя нейтронов.
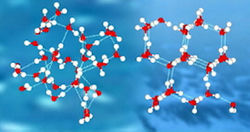
Вода – уникальный материал для хранения и передачи информации. Новые результаты исследования структуры воды, полученные на сей раз учеными из США, подтверждают наличие в ней крупномасштабной структуры из ансамблей молекул. Ранее об этом говорили представители научного центра прикладных исследований (НЦПИ) Объединенного института ядерных исследований в подмосковной Дубне.
Новое исследование, проведенное учеными национальной лаборатории Лоуренс-Беркли министерства энергетики США, во многом подтверждает предположение подмосковных ученых о наличии в воде сверхмолекулярной структуры и о ее особенностях. Им удалось показать, что молекулы воды способны за счет водородных связей образовывать устойчивые образования – истинные «кирпичики» воды, представляющие собой топологически цепочки и кольца из множества молекул.
Научно исследовательский институт биологии при Иркутском государственном университете (Иркутск)
Иркутский государственный университет (Иркутск)
Исследовалось влияние воды с разным содержанием дейтерия дождевых червей. Дождевые черви выбирали садки с более легкой водой (13 ppm) и избегали воду, обогащенную дейтерием. В более лег кой воде масса червей увеличивается быстрее, чем в тяжелой.
Питьевая вода – это сложный по своей изотопной структуре и составу продукт, оказывающий полифизиологическое действие на организм человека. На долю лёгкой (протиевой воды), состоящей из обычных лёгких изотопов водорода 1Н и кислорода 16О приходится примерно 99,7% всех атомов изотопов. Остальную долю составляют тяжелые изотопы - H218О (73,5%), H217О (14,7%) и HD16О (11,5%). В воде пресноводных источников содержание тяжелой воды колеблется от 150 до 300 мг/л (в расчете на молекулу HDO), а тяжелокислородной (Н218О) – около 2 г/л. Это сопоставимо или даже превышает допустимое содержание солей в питьевой воде. Несмотря на этот факт, влияние примесей тяжелых молекул воды на потребительские свойства питьевой воды изучено недостаточно.
Обычная питьевая вода только на 99,7% состоит из легкой воды, молекулы которой образованы легкими атомами водорода и кислорода. В виде примеси в любой природной воде присутствует и тяжёлая вода, которая в чистом виде является ядом для всего живого.
Тяжёлая вода (оксид дейтерия) — имеет ту же химическую формулу, что и обычная вода, но вместо атомов водорода содержит два тяжёлых изотопа водорода — атомы дейтерия. Внешне тяжёлая вода выглядит как обычная — бесцветная жидкость без вкуса и запаха, а вот по своим физико-химическим свойствам и негативному воздействию на организм тяжёлая вода сильно отличается от лёгкой воды.
Доклад к.х.н. А.А. Тимакова на 8-й Всероссийской (международной) научной конференции ФИЗИКО – ХИМИЧЕСКИЕ ПРОЦЕССЫ ПРИ СЕЛЕКЦИИ АТОМОВ И МОЛЕКУЛ
Генеральный директор компании Лангвэй кандидат химических наук Александр Алексеевич Тимаков (на фото).
Вступление. Рад снова оказаться среди людей, которых не пугает слово «изотоп» и которые не вздрагивают при упоминании о «тяжелой воде». Но даже здесь необходимо сразу пояснить термин «легкая вода». Под легкой водой далее мы подразумеваем воду, содержание дейтерия и кислорода-18 в которой снижено по отношению к природному уровню, а, точнее, к стандартной среднеокеанической воде (SMOW), потому что природный уровень, как мы покажем далее, понятие растяжимое, особенно в отношении дейтерия.
Лёгкая вода «Лангвей» рекомендуется для быстрой и глубокой очистки организма, что необходимо при нарушениях обменных процессов, перед операцией и в послеоперационный период, а также при лечении опухолевых заболеваний.
У человека повышается физическая активность, появляется дополнительная энергия, отступают болезни, замедляется процесс старения.
Легкая вода «Лангвей» незаменима при: повышенном артериальном давлении; сахарном диабете; избыточной массе тела.
Лёгкая вода – это сложный по своей структуре и составу продукт, оказывающий полифизиологическое действие на организм человека. В этой связи важно оценить, какое влияние на организм окажет очистка питьевой воды от тяжелых молекул при сохранении всех других компонентов воды на регламентируемых гигиеническими нормативами уровнях. Учитывая роль воды в организме и известные изотопные эффекты тяжелой воды, и результаты, полученные по легкой воде, можно ожидать, что наибольший эффект такая очистка может оказать на свойства биологических мембран, регуляторные системы и энергетический аппарат живой клетки. Хорошо известно, например, что под влиянием тяжелой воды ингибируется инициируемый глюкозой выход инсулина из ткани поджелудочной железы и островков Лангерганса, уменьшается скорость поглощения кислорода митохондриями клеток.