ADAPTATION TO 2H2O
AND BIOPHYSICAL PROPERTIES OF [U -2H]LABELED MACROMOLECULES
FIGURE
The imaginary principle of realization of biological adaptation
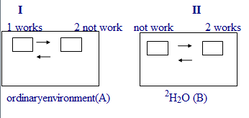
4.1. The main hypothese. We proposed that a cell theoretically could in principle synthezise a big number of forms of [2H]labeled macromolecules with somewhat different structures and conformations, so that a cell could easily select a preferable one from al these species in a course of adaptation to 2H2O, that is the best suitable namely for that conditions. A simple imaginary principle I am going to discuss here perhaps somewhat may explain this probable mechanism. Let us suppose, for example that there are at least two imadinary structural systems - ordinary (normal) system call it a system 1 and unordinary (adaptive) system 2 (see a Figure above). Supporse, that the environment is a homoginious substanse and compose from ordinary substance A (H2O) (situation 1). The necessarely condition for the normal working of this model in natural H2O environment is that system 1 works and system 2 stay in background (situation 2). Supporse, that the environment have changed for substance B (2H2O). Then the system 2 will work, while the system 1 will stay in background (situation 2). When environment will be the natural again, the system 1 will begin the work again, while the system 2 will stay in background. Admitt, that the two systems both presented at the time being and could be regulated in such way that they may switch bitween each other during the working so that the model system does not undergoing the considerable alterations.
4.2. Phenomenon of biological adaptation to 2H2O. Our research has confirmed, that ability to adaptation to 2Н2О is differed for various species of bacteria and can to be varried even in frames of one taxonomic family (Mosin O. V. et al., 1996a, 1996b).From this, it is possible to conclude, that the adaptation to 2Н2О is determined both by taxonomic specifity of the organism, and peculiarities of the metabolism, as well as by functioning of various ways of accimilation of hydrogen (deuterium) substrates, as well as evolutionary level, which an object itself occupies. The less a level of evolutionary development of an organism, the better it therefore adapts itself to 2H2O. For example, there are halophilic bacteria that are being the most primitive in the evolutionary plan, and therefore, they practically not requiring to carry out a special adaptation methods to grow on 2Н2О. On the contrary, bacills (eubacteria) and methylotrophs (gram-negative bacteria) worse adapted to 2Н2О. At the same time for all tested cells the growth on 2H2O was accompanied by considerable decrease of a level of biosynthesis of appropriated cellular compounds. The data obtained confirm that the adaptation to 2Н2О is a rather phenotypical phenomenon, as the adapted cells could be returned to a normal growth and biosynthesis in protonated media after lag-phase (Mosin O. V. et al., 1993). However, when the adaptive process goes continuously during the many generation, the population of cells can use a special genetic mechanisms for the adaptation to 2H2O. For example, mutations of geens can be resulted in amino acid replacements in molecules of proteins, which in turn could cause a formation of a new isoenzymes, and in the special cases - even the anomal working enzymes of a newer structure type. The replacements of these compounds can ensure a development of new ways of regulation of enzymic activity, ensuring more adequate reaction to signals, causing a possible changes in speeds and specifity of metabolic processes. Despite it, the basic reactions of metabolism of adapted cells probably do not undergo essential changes in 2Н2О. At the same time the effect of convertibility of growth on Н2О/2Н2О - does not theoretically exclude an opportunity that this attribute is stably kept when cells grown on 2Н2О, but masks when transfer the cells on deuterated medium. However, here it is necessary to emphasize, that for realization of biological adaptation to 2H2O the composition of growth medium plays an important role. In this case it is not excluded, that during the adaptation on the minimal medium, containing 2Н2О there are formed the forms of bacteria, auxotrophic on a certain growth factors (for example amino acids et) and thereof bacterial growth is inhibited while grown on these media. At the same time the adaptation to 2Н2О occurs best on complex media, the composition of which coul compensate the requirement in those growth factors. It is possible also to assume, that the macromolecules realize the special mechanisms, which promote a stabilization of their structure in 2H2O and the functional reorganization for best working in 2Н2О. Thus, the distinctions in nuclear mass of hydrogen atom and deuterium can indirectly to be a reason of distinctions in synthesis of deuterated forms of DNA and proteins, which can be resulting in the structural distinctions and, hence, to functional changes of [2H]labeled macromolecules. Hawever, it is not excluded, that during incubation on 2Н2О the enzymes do not stop the function, but changes stipulating by isotopic replacement due to the primary and secondary isotopic effects as well as by the action of 2Н2О as solvent (density, viscosity) in comparison with Н2О are resulted in changes of speeds and specifics of metabolic reactions.
In the case with biological adaptation to 2H2O we should inspect the following types of adaptive mechanisms: 1. adaptation at a level of macromolecular components of cells: It is possible to allocate mainly two kinds of such adaptation: (a). Differences of intracellular concentration of macromolecules; (b). The forming in 2H2O the deuterated macromolecules with other conformations, which could be replaced the ordinary protonated macromolecules synthesized by cells in normal conditions. We suppose, that in principle, any protein macromolecule could adopt an almost unlimited number of conformations. Most pilypeptide chains, however, fold into only one particular conformation determined by their amino acid sequence. That is because the side chains of the amino acids associate with one another and with water (2H2O) to form various weak noncovalent bonds. Provided that the appropriate side chains are present at crucial positions in the chain, large forces are developed that make one particular conformation especially stable.
These two strategies of adaptation could possible to be distinqueshed accordinly as "quantitative" and "qualitative" strategies; 2. adaptation at a level of microenvironment in wich macromolecules are submerged: the essence of this mechanism is, that the adaptive change of structural and conformational properties of [2H]labeled macromolecules is conditioned both by directional action of 2H2O environment on a growth of cells and by its physico-chemical structure (osmotic pressure, viscosity, density, рН, concentration of 2H2O). 2H2O appeared to stabilize the plasmagel structure of biological microenvironment. The external pressure required to make the cells assume a spherical shape increased 3.6 kg/cm2 for each per cent increase in the presence of 2H2O. It thus seems well established that deuteration can affect the mechanical properties of cytoplasm, and that this factor must be taken into account in assessing the consequences of isotopic substitution of macromolecules. In model experiments with gelatin structure, it was demonstrated that in 2H2O there is a greater protein-protein interaction than in H2O (Scheraga J. A; 1960). A progressive increase in the melting temperature of the gel in 2H2O is observed accompanied by an increase in the reduced viscosity. That 2H2O can have marked effects on the physical properties of proteins has been known for some time. Consequently it is natural to attribute changes in the mechanical properties of cell structures induced by 2H2O to protein response. Nevertheless, the effects of deuterium on proteins, while real, must be only a partial explanation of the situation. The interaction of proteins with solvent water is extraordinarily complex, and the exact nature of the protein is crucial in determining the magnitude of changes resulting from the replacement of H2O by 2H2O.
This mechanism has extremely large importance and supplements the macromolecular adaptation; 3. adaptation at a functional level, when the change of an overall performance of macromolecular systems, is not connected with a change of a number of macromolecules being available or with the macromolecules of their types. Adaptation in this case could provide the changes by using the already existing macromolecular systems - according to requirements by this or that metabolic activity.
TABLE
Some physical constants of ordinary and heavy water
Physical constant
H2O
2Н2О
Density, d20 (g/c.c)
0,9982
1,1056
Molecular volume, V20 (ml/mole)
18,05
18,12
Viscosity m20 (centipose)
1,005
1,25
Melting point (0C)
0,1
3,82
Boiling point (0C)
100,0
101,72
Temperature of maximum density (0C)
4,0
11,6
Ion product (25 0C)
10-14
0,3x10-14
Heat of formation (cal/mole)
-68,318
-70,414
Free energy of formation (cal/mole)
-56,693
-58,201
Entropy (e.u/mole)
45,14
47,41
Secondary effects may still be of importance in biological systems sensitive to kinetic distortions. Deuterium also affects equilibrium constants, particularly the ionization constants of weak acids and bases in composition of macromolecules dissolved in heavy water (see a Table below). Acid strength of macromolecules in 2H2O is decreased by factors of 2 to 5, and consequently, the rates of acid-base catalyzed reactions may be greatly different in 2H2O as compared to H2O. Such reactions frequently may be a faster in 2H2O than H2O solution (Covington A. K., Robinson R. A., and Bates R. G., 1966; Glasoe P. K., and Long F. A., 1960).
4.2. The chemical isotopic effect of 2H2O. The effect of isotopic replacement that has particularly attracted the attention of chemists is the kinetic isotope effect (Thomson J. F., 1963). The substitution of deuterium for hydrogen in a chemical bond of macromolecules can markedly affect the rate of scission of this bond, and so exert pronounced effects on the relative rates of chemical reactions going in 2H2O with participation of macromolecules. This change in rate of scission of a bond resulting from the substitution of deuterium for hydrogen is a primary isotopic effect. The direction and magnitude of the isotope effect will depend on the kind of transition state involved in the activated reaction complex, but in general, deuterium depresses reaction rates. The usual terminology of the chemist to describe the primary kinetic effect is in terms of the ratio of the specific rate constants kh/kd. The maximum positive primary kinetic isotopic effect which can be expected at ordinary temperatures in a chemical reaction leading to rupture of bonds involving hydrogen can be readily calculated, and the maximum ratio kh/kd in macromolecules is in the range of 7 to 10 for C-H versus C-2H, N-H versus N-2H, and O-H versus O-2H bonds. However, maximum ratios are seldom observed for a variety of reasons, but values of kh/kd in the range of 2 to 5 are common (Wiberg K. B., 1955). Deuterium located at positions in a macromolecule other than at the reaction locus can also affect the rate of a reaction. Such an effect is a secondary isotope effect and is usually much smaller than a primary isotope effect.
In general, when the macromolecules transfer to deuterated medium not only water due to the reaction of an exchange (Н2О -2Н2О) dilutes with deuterium, but also occurs a very fast isotopic (1Н-2Н)-exchange in hydroxylic (-OH), carboxilic (-COOH), sulfurhydrilic (-SH) and nitrogen (-NH; -NH2) groups of all organic compounds including the nucleic acids and proteins. It is known, that in these conditions only С-2Н bond is not exposed to isotopic exchange and thereof only the species of macromolecules with С-2H type of bonds can be synthesized de novo. This is very probably, that the most effects, observed at adaptation to 2Н2О are connected with the formation in 2Н2О [U -2H]labeled molecules with conformations having the other structural and dynamic properties, than conformations, formed with participation of hydrogen, and consequently having other activity and biophysical properties. So, according to the theory of absolute speeds the break of С-1H-bonds can occur faster, than С-2H-bonds (C-2H-bonds are more durable than C-1 , mobility of an ion 2H+ is less, than mobility of 1Н+, the constant of ionization 2Н2О is a little bit less than ionization constant of 2Н2О. Thus, in principle, the structures of [U -2H]labeled macromolecules may to be more friable that those are forming in ordinary H2O. But, nevertheless, the stability of [U -2H]labeled macromolecules probably depending on what particular bond is labeled with deuterium (covalent bonds -C2H that causing the instability or hydrogen bonds causing the stabilization of conformation of macromolecules via forming the three-dimentional netwok of hydrogen(deuterum) bonds in macromolecule) and what precise position of the macromolecule was labeled with deuterium. For example, the very valuable and sensitive for deuterium substitution position in macromolecule is the reactive center (primary isotopic effects). The non-essential positions in macromolecule are those ones that situated far away from the reactive center of macromolecule (secondary isotopic effects). It is also possible to make a conclusion, that the sensitivity of various macromolecules to substitution on 2Н bears the individual character and depending on the structure of macromolecule itself, and thus, can be varried. From the point of view of physical chemistry, the most sensitive to replacement of 1Н+ on 2H+ can appear the apparatus of macromolecular biosyntesis and respiration system, those ones, which use high mobility of protons (deuterons) and high speed of break of hydrogen (deuterium) bonds. From that it is posible to assume, that the macromolecules should realize a special mechanisms (both at a level of primary structure and a folding of macromolecules) which could promote the stabilizition of the macromolecular structure in 2H2O and somewhat the functional reorganization of their work in 2H2O. A principal feature of the structure of such biologically important compounds as proteins and nucleic acids is the maintenance of their structure by virtue of the participation of many hydrogen bonds in macromolecule. One may expect that the hydrogen bonds formed by of many deuterium will be different in their energy from those formed by proton. The differences in the nuclear mass of hydrogen and deuterium may possibly cause disturbances in the DNA-synthesis, leading to permanent changes in its structure and consequently in the cells genotype. The multiplication which would occur in macromolecules of even a small difference between a proton and a deuteron bond would certainly have the effect upon its structure. The sensitivity of enzyme function to structure and the presumed sensitivity of nucleic acids function (genetic and mitotic) to its structure would lead one to expect a noticeable effect on the metabolic pattern and reproductive behavior of the organism. And next, the changes in dissociation constants of DNA and protein ionizable groups when transfer the macromolecule from water to 2H2O may perturb the charge state of the DNA and protein. Substitution of 1H for deuterium also affects the stability and geometry of hydrogen bonds in apparently rather complex way and may, through the changes in the hydrogen bond zero-point vibrational energies, alter the conformational dynamics of hydrogen (deuterium)-bonded structures within the DNA and protein in 2H2O.